
羟基酪醇、酪醇及其衍生物及其对人类健康的潜在影响



抽象的
1. 简介
2. 方法论
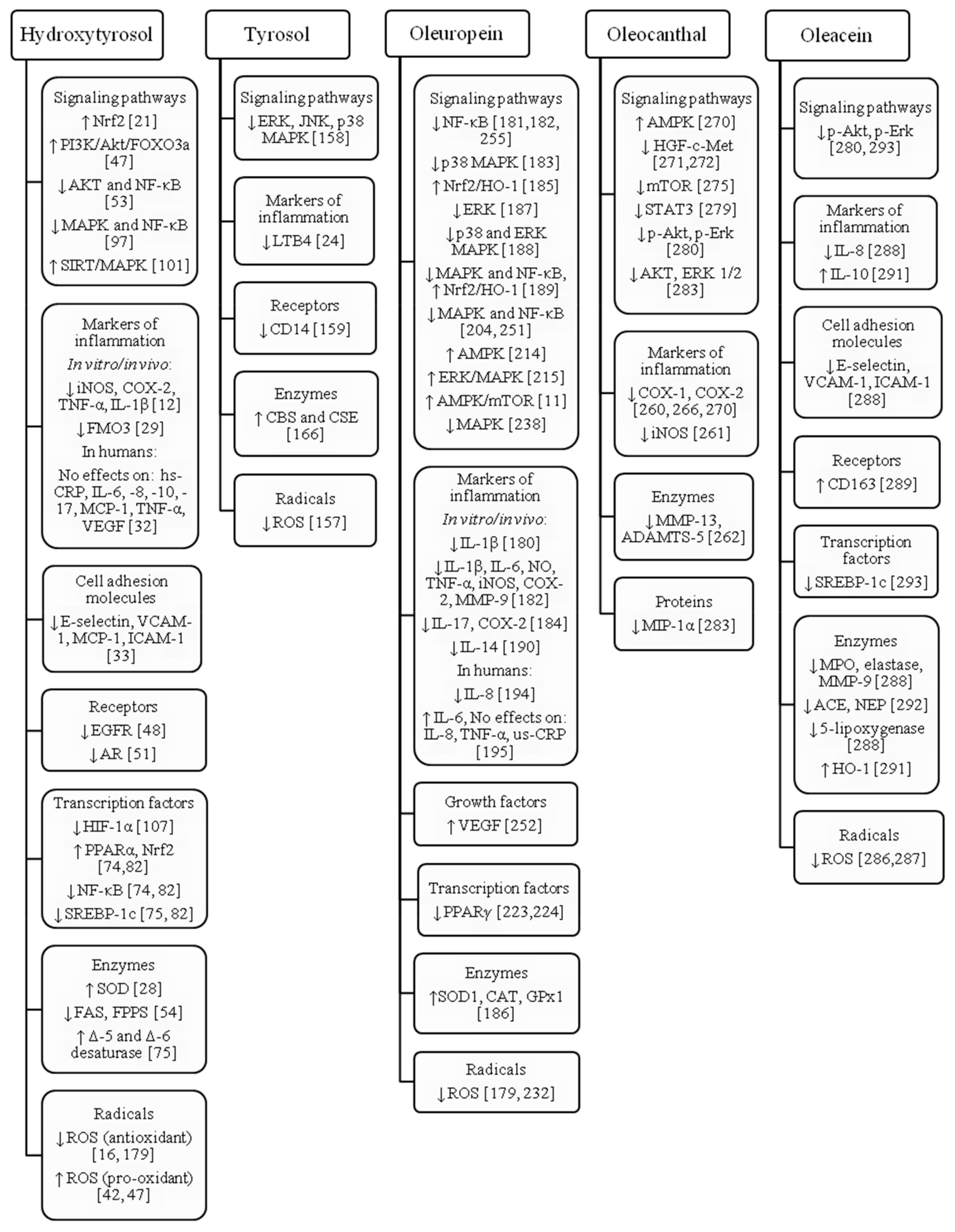
3.羟基酪醇


3.1. 抗动脉粥样硬化和心脏保护作用
3.2. 抗癌作用
3.3. 神经保护作用
3.4. 抗糖尿病、调脂和减肥作用
3.5. 抗菌和抗病毒作用
3.6. 其他影响
3.7. 新陈代谢


4. 酪醇
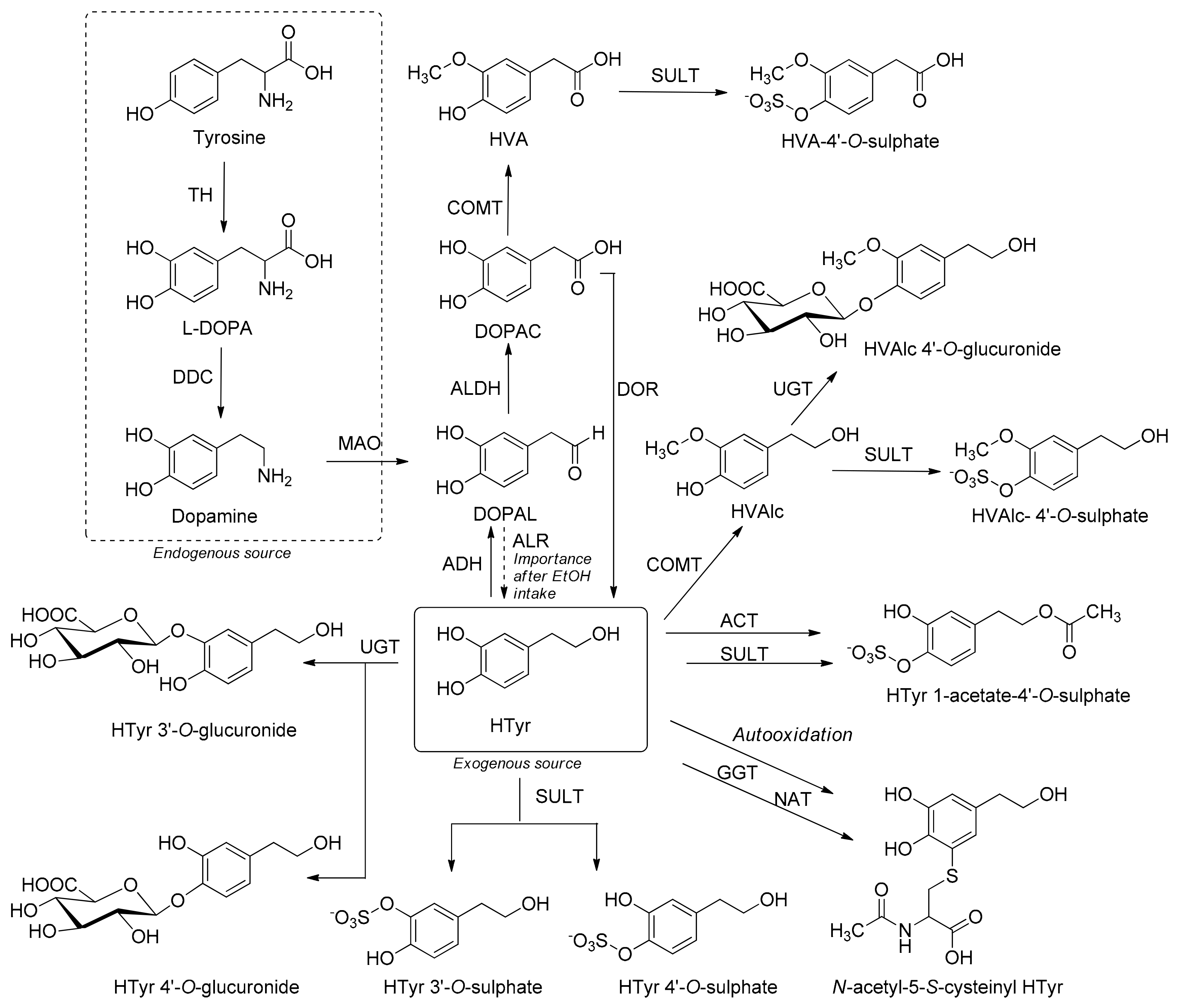
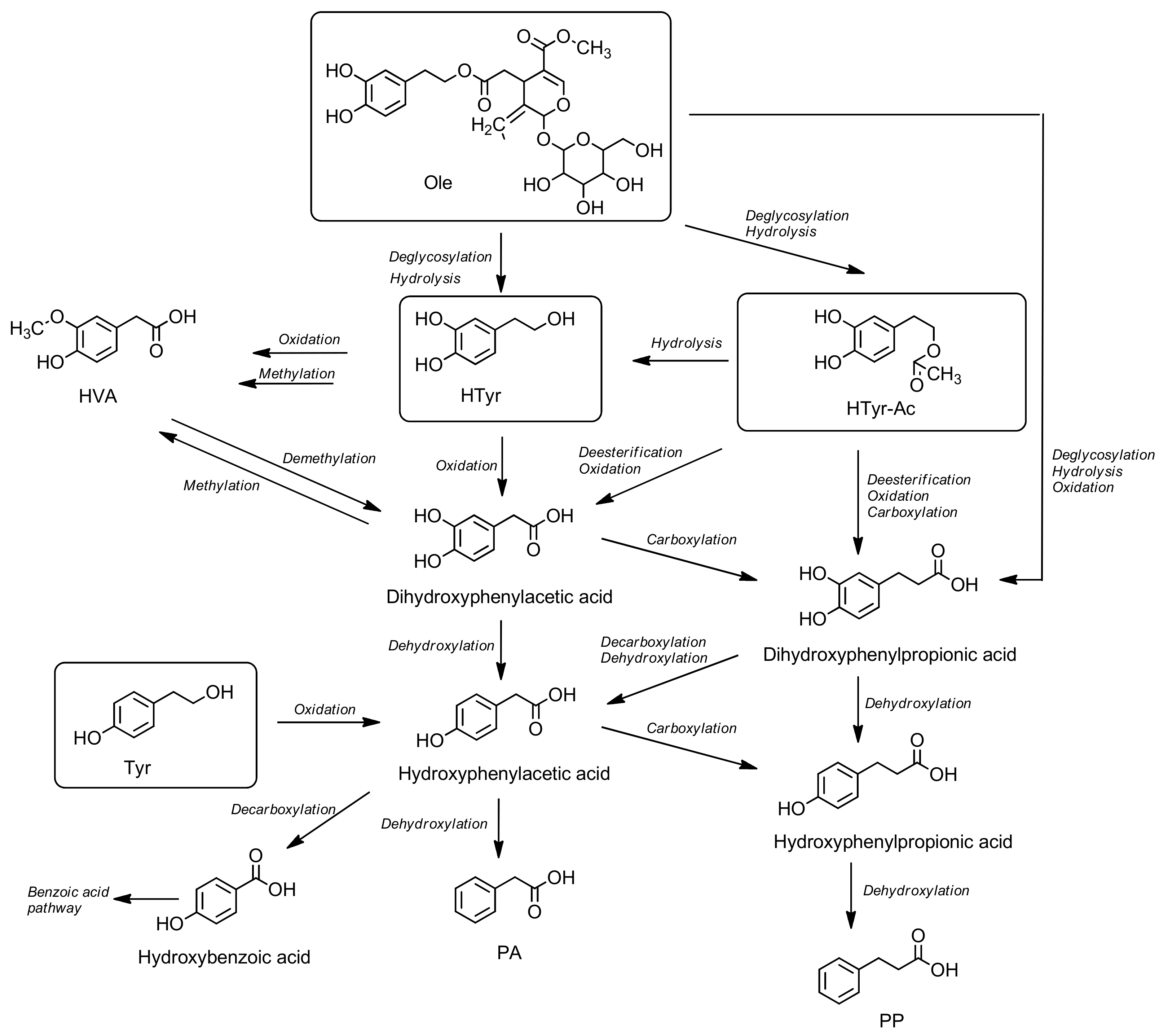
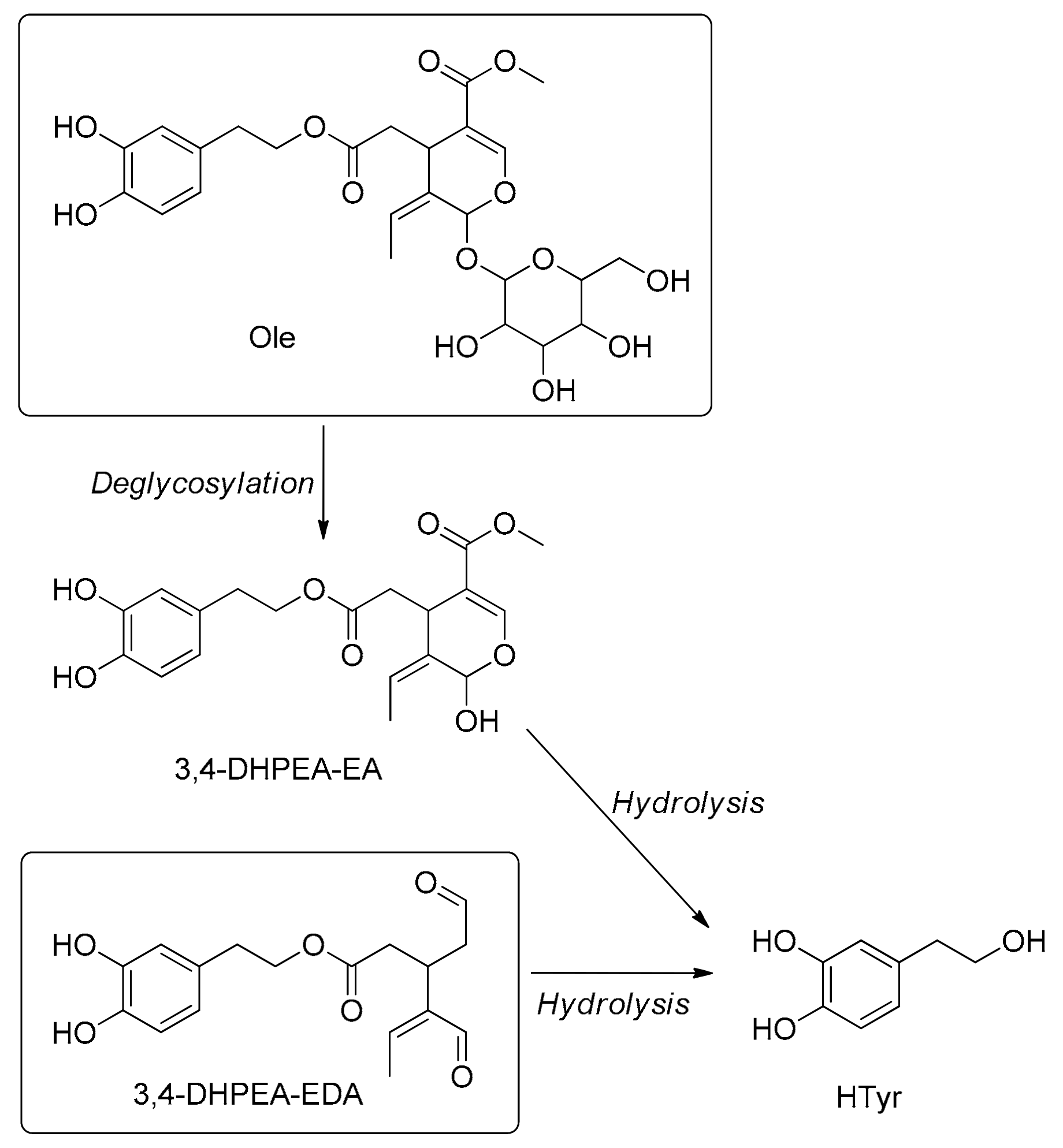
代谢

5.橄榄苦苷
5.1. 抗动脉粥样硬化和心脏保护作用
5.2. 抗癌作用
5.3. 抗糖尿病、调脂和抗肥胖作用
5.4. 神经保护作用
5.5. 抗菌和抗病毒作用
5.6. 骨质保护作用
5.7. 其他影响
5.8. 新陈代谢

6. 油橄榄刺激醛
7. 橄榄素
8. 潜在的临床应用
9. 结论
作者贡献
资金
致谢
利益冲突
参考
- 洛佩斯-米兰达,J.;佩雷斯·希门尼斯,F.;玫瑰。;德卡特琳娜,R.;巴迪蒙,L.;密歇根州科瓦斯;埃斯克里奇,E.;奥尔多瓦斯,JM;索里格,F.;阿比亚,R.;等人。橄榄油与健康:第二届橄榄油与健康国际会议共识报告摘要,哈恩和科尔多瓦(西班牙),2008 年。元数据。心血管。迪斯。 2010 , 20 , 284–294。 [谷歌学术] [交叉引用] [ PubMed ]
- 埃斯特鲁奇,R.;玫瑰。;萨拉斯-萨尔瓦多,J.;科瓦斯,M.-I.;科雷拉,D.;阿罗斯,F.;戈麦斯-格拉西亚,E.;鲁伊斯-古铁雷斯,V.;菲奥尔,M.;拉佩特拉,J.;等人。通过补充特级初榨橄榄油或坚果的地中海饮食对心血管疾病进行一级预防。N. 英格兰。 J. Med。 2018,378,e34。 [谷歌学术] [交叉引用] [ PubMed ]
- Sofi, F.;Macchi, C.;Abbate, R.;Gensini, GF;Casini, A. 地中海饮食与健康状况:最新的荟萃分析和基于文献的依从性评分提案。公共卫生营养。2014 年,17,2769–2782。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Covas, M.-I.;de la Torre, R.;Fitó, M. 初榨橄榄油:预防心血管风险的关键食品。英国营养学杂志。2015 年,113,S19–S28。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Psaltopoulou, T.;Kosti, RI;Haidopoulos, D.;Dimopoulos, M.;Panagiotakos, DB 橄榄油摄入量与癌症患病率呈负相关:对 19 项观察性研究中的 13800 名患者和 23340 名对照者的系统评价和荟萃分析。脂质健康疾病。2011 年,10,127–143。[ Google 学术搜索] [ CrossRef ]
- Buckland, G.;Gonzalez, CA 橄榄油在疾病预防中的作用:重点关注队列研究和饮食干预试验的最新流行病学证据。英国营养学杂志。2015 年,113,S94–S101。[ Google 学术搜索] [ CrossRef ]
- 皮罗迪,M.;阿尔比尼,A.;法比亚尼,R.;乔瓦内利,L.;卢塞里,C.;纳特拉,F.;罗西诺利,P.;罗西,T.;塔蒂奇,A.;塞尔维利,M.;等人。特级初榨橄榄油的营养基因组学:综述。生物因素 2017 , 43 , 17–41。 [谷歌学术] [交叉引用] [ PubMed ]
- 梅嫩德斯,JA;乔文,J.;阿拉贡内斯,G.;巴拉洪-加泰罗尼亚,E.;贝尔特兰-德邦,R.;博拉斯-利纳雷斯,I.;坎普斯,J.;科罗米纳斯-法贾,B.;库菲,S.;费尔南德斯-阿罗约,S.;等人。特级初榨橄榄油中环烯醚萜多酚的异种激素和抗衰老活性:一个新的衰老抑制剂家族。细胞 周期2013,12,555–578。 [谷歌学术] [交叉引用]
- Ghanbari, R.;Anwar, F.;Alkharfy, KM;Gilani, A.-H.;Saari, N. 橄榄 (Olea europaea L.) 不同部位的宝贵营养成分和功能性生物活性物质——综述。国际分子科学杂志。2012 年, 13,3291–3340 。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Rodriguez - Morato,J. ; Xicota,L.;Fito,M. ; Farre,M.;Dierssen,M.;de la Torre,R. 橄榄油酚类化合物在预防神经退行性疾病中的潜在作用。Molecules 2015,20,4655–4680。[ Google Scholar ] [ CrossRef ] [绿色版本]
- Rigacci,S. ; Stefani,M. 橄榄油多酚的营养特性。从培养细胞到动物模型再到人类的路线图。Int . J. Mol. Sci. 2016,17,843。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- Parkinson, L.;Cicerale, S. 初榨橄榄油酚类化合物的健康益处机制。Molecules 2016,21,1734 。 [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jakobušić Brala, C.;Barbarić, M.;Karković Marković, A.;Uršić, S。橄榄油酚类化合物的生物医学方面和活性。《橄榄油手册。酚类化合物、生产和健康益处》,第 1 版;Miloš, J.,编辑;Nova Science Publishers:美国纽约州纽约,2017 年;第 47-85 页。ISBN 978-1-53612-356-2。[ Google 学术]
- Cicerale,S.;Lucas, L .;Keast,R. 特级初榨橄榄油中的抗菌、抗氧化和抗炎酚类活性。Curr . Opin. Biotech。2012,23,129–135 。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- Fabiani,R .橄榄油裂环烯醚萜酚的抗癌特性:体内研究的系统评价。食品功能 。2016,7,4145–4159。[ Google学术搜索] [ CrossRef ] [ PubMed ]
- Visioli,F.;Poli , A.;Galli,C. 橄榄和橄榄油中酚类物质的抗氧化和其他生物活性。Med . Res. Rev. 2002,22,65–75 。 [ Google Scholar ] [ CrossRef ] [ PubMed ]
- 托梅-卡内罗,J.;克雷斯波,MC;伊格莱西亚斯-古铁雷斯,E.;马丁,R。吉尔-萨莫拉诺,J.;托马斯-扎皮科,C.;布尔戈斯-拉莫斯,E.;科雷亚,C.;戈麦斯-科罗纳多,D.;马萨诸塞州拉松西翁;等人。羟基酪醇补充剂可调节啮齿动物和人类中 miRNA 的表达。J.努特尔。生物化学。 2016 , 34 , 146–155。 [谷歌学术] [交叉引用]
- Echeverria, F.;Ortiz, M.;Valenzuela, R.;Videla, LA 羟基酪醇和细胞保护:临床干预的预测。国际分子科学杂志。2017 年,18,930。[ Google 学术搜索] [ CrossRef ]
- Robles-Almazan,M.;Pulido-Moran,M.;Moreno-Fernandez,J.;Ramirez-Tortosa,C.;Rodriguez-Garcia,C.;Quiles,JL;Ramirez-Tortosa,MC。羟基酪醇:生物利用度、毒性和临床应用。食品研究国际版 2018,105,654-667。[ Google 学术搜索] [ CrossRef ]
- Wani, TA;Masoodi, FA;Gani, A.;Baba, WN;Rahmanian, N.;Akhter, R.;Wani, IA;Ahmad, M. 橄榄油及其主要生物活性化合物:羟基酪醇——近期文献综述。食品科学技术趋势 。2018年,77,77–90。[ Google 学术搜索] [ CrossRef ]
- Bayram, B.;Ozcelik, B.;Grimm, S.;Roeder, T.;Schrader, C.;Ernst, IMA;Wagner, AE;Grune, T.;Frank, J.;Rimbach, G. 富含橄榄油酚类物质的饮食通过诱导 Nrf2 依赖性基因表达降低 SAMP8 小鼠心脏的氧化应激。Rejuvenation Res。2012 年,15,71-81。[ Google 学术搜索] [ CrossRef ] [绿色版本]
- Scoditti,E.;Nestola,A.;Massaro,M.;Calabriso,N.;Storelli,C.;De Caterina,R.;Carluccio,MA 羟基酪醇通过抑制 PKCα 和 PKCβ1 抑制活化人单核细胞中的 MMP-9 和 COX-2 活性和表达。动脉粥样硬化 2014,232,17-24。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Bigagli,E.;Cinci,L.;Paccosi,S.;Parenti,A.;D'Ambrosio,M.;Luceri,C. 营养相关浓度的白藜芦醇和羟基酪醇可减轻人类粒细胞和单核细胞的氧化爆发以及 LPS 刺激的 RAW 264.7 巨噬细胞中促炎介质的产生。国际免疫药理学。2017 年,43,147-155。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Perona, JS;Cabello-Moruno, R.;Ruiz-Gutierrez, V. 初榨橄榄油成分在调节内皮功能中的作用。J . Nutr. Biochem. 2006 , 17 , 429–445。[ Google Scholar ] [ CrossRef ] [绿色版本]
- Rietjens, SJ;Bast, A.;Haenen, GRMM 对橄榄油抗氧化剂羟基酪醇抗氧化潜力争议的新见解。农业与食品化学杂志。2007 年,55,7609–7614。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- EFSA饮食产品、营养和过敏( NDA) 专家组。关于维生素 E 和保护 DNA、蛋白质和脂质免受氧化损伤的健康声明的科学意见。EFSA J. 2010,8,1816。[ Google Scholar ] [ CrossRef ]
- Berrougui,H.;Ikhlef,S.;Khalil,A. 特级初榨橄榄油多酚促进胆固醇流出并改善 HDL 功能。基于证据的补充。替代医学。2015 年,2015 年,208062。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Calabriso,N.;Gnoni,A.;Stanca,E.;Cavallo,A.;Damiano,F.;Siculella,L.;Carluccio,MA 羟基酪醇通过预防线粒体功能障碍改善炎症条件下的内皮功能。氧化医学细胞长寿 。2018,2018,9086947 。 [ Google学术] [ CrossRef ] [ PubMed ]
- Wu, X.; Li, C.; Mariyam, Z.; Jiang, P.; Zhou, M.; Zeb, F.; ul Haq, I.; Chen, A.; Feng, Q. 丙烯醛通过刺激含有单加氧酶 3 的肝黄素和保护羟基酪醇诱导动脉粥样硬化形成。J. Cell. Physiol. 2019 , 234 , 475–485。[ Google 学术搜索] [ CrossRef ]
- Fuccelli,R.;Fabiani ,R .;Rosignoli,P. 羟基酪醇在小鼠全身炎症模型中发挥抗炎和抗氧化作用。Molecules 2018,23,3212。[ Google Scholar ] [ CrossRef ]
- 科利卡,C.;迪伦佐,L.;特罗姆贝塔,D.;斯梅里利奥,A.;贝尔纳迪尼,S.;乔科洛尼,G.;米兰达科斯塔,R.;瓜尔蒂里,P.;西尼巴尔迪·萨利梅,P.; De Lorenzo, A. 基于羟基酪醇的药物制剂对身体成分、代谢状态和基因表达的抗氧化作用:随机双盲、安慰剂对照交叉试验。氧化物。医学。细胞。长寿。 2017 , 2473495. [谷歌学术] [交叉引用]
- Crespo, MC;Tome-Carneiro, J.;Burgos-Ramos, E.;Kohen, VL;Espinosa, MI;Herranz, J.;Visioli, F. 给人类服用羟基酪醇一周不会激活 II 期酶。药理学研究 。2015 年,95–96,132–137。[ Google 学术搜索] [ CrossRef ]
- Catalan, U.; Lopez de las Hazas, M.-C.; Pinol, C.; Rubio, L.; Motilva, M.-J.; Fernandez-Castillejo, S.; Sola, R. 羟基酪醇及其主要血浆循环代谢物通过抑制 MAPK 通路减弱动脉粥样硬化的初始步骤。J . Funct. Food 2018,40,280-291 。 [ Google Scholar ] [ CrossRef ]
- Gonzalez-Santiago,M.;Martin-Bautista,E.;Carrero,JJ;Fonolla,J.;Baro,L.;Bartolome,MV;Gil-Loyzaga,P.;Lopez-Huertas,E. 给高脂血症兔子服用羟基酪醇(一种存在于橄榄油中的酚类抗氧化剂)一个月,可以改善血脂状况、抗氧化状态并减少动脉粥样硬化的发展。动脉粥样硬化 2006,188,35-42 。 [ Google 学术搜索] [ CrossRef ]
- Tomé- Carneiro,J.;Visioli,F. 基于多酚的营养品用于预防和治疗心血管疾病:人类证据回顾。植物医学 2016,23,1145–1174。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Lopez-Huertas,E.;Fonolla,J. 羟基酪醇补充剂可增加体内维生素 C 水平。一项人类志愿者试验。氧化还原。生物学 。2017 年,11,384-389。[ Google 学术搜索] [ CrossRef ]
- Xie, Y.; Xu, Y.; Chen, Z.; Lu, W.; Li, N.; Wang, Q.; Shao, L.; Li, Y.; Yang, G.; Bian, X. 一种具有抗糖尿病、抗高血脂、抗氧化和抗炎作用的新型多功能羟基酪醇-非诺贝特。生物医学。药物治疗学。2017,95,1749–1758 。 [ Google Scholar ] [ CrossRef ] [ PubMed ]
- 谢永东;陈志忠;李娜;陆文峰;徐永华;林燕燕;邵玲华;王庆天;郭林燕;高永强等。羟基酪醇烟酸酯是一种新型多功能降血脂降血糖剂。生物医学药物治疗学。2018,99,715–724 。[ Google学术搜索] [ CrossRef ]
- Xie, Y.-D.; Chen, Z.-Z.; Shao, L.-H.; Wang, Q.-T.; Li, N.; Lu, W.-F.; Xu, Y.-H.; Gao, Y.-Q.; Guo, L.-Y.; Li, Y.-P.; et al. 一种具有降血脂、抗氧化和保肝作用的新型多功能羟基酪醇-氯贝特。Bioorg . Med. Chem. Lett. 2018 , 28 , 3119–3122。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- Gonzalez-Correa,JA;Navas,MD;Munoz-Marin,J.;Trujillo,M.;Fernandez-Bolanos,J.;Pedro de la Cruz,J. 与乙酰水杨酸相比,给大鼠施用羟基酪醇和羟基酪醇乙酸酯对血小板功能的影响。农业。食品化学杂志。2008,56,7872-7876 。 [ Google学术] [ CrossRef ]
- Catalan, U.; Rubio, L.; Lopez de las Hazas, M.-C.; Herrero, P.; Nadal, P.; Canela, N.; Pedret, A.; Motilva, M.-J.; Sola, R. 羟基酪醇及其复合形式 (secoiridoids) 调节健康大鼠的主动脉和心脏蛋白质组:潜在的心脏保护作用。Mol . Nutr. Food Res. 2016 , 60 , 2114–2129. [ Google Scholar ] [ CrossRef ] [ PubMed ] [绿色版本]
- Fabiani,R.;Sepporta,MV;Rosignoli,P.;De Bartolomeo,A.;Crescimanno,M.;Morozzi,G. 羟基酪醇对不同肿瘤细胞的抗增殖和促凋亡活性:细胞外过氧化氢产生的作用。欧洲营养学杂志。2012,51,455–464 。 [ Google 学术搜索] [ CrossRef ]
- Imran, M.; Nadeem, M.; Gilani, SA; Khan, S.; Sajid, MW; Amir, RM 橄榄苦苷及其代谢物羟基酪醇的抗肿瘤观点:最新更新。食品科学杂志。2018 , 83 , 1781–1791。[ Google Scholar ] [ CrossRef ] [ PubMed ] [绿色版本]
- Corona, G.;Tzounis, X.;Assunta DessÌ, M.;Deiana, M.;Debnam, ES;Visioli, F.;Spencer, JPE 橄榄油多酚在胃肠道中的命运:胃和结肠微生物群依赖性生物转化的影响。自由基研究 。2006 年,40,647–658。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Corona,G.;Deiana,M.;Incani,A.;Vauzour,D.;Dessi,MA;Spencer,JPE 羟基酪醇通过抑制 ERK1/2 和细胞周期蛋白 D1 来抑制人类结肠腺癌细胞的增殖。分子营养食品研究。2009 年,53,897-903。[ Google 学术搜索] [ CrossRef ]
- De las Hazas,MCL;Piñol,C. ; Macià , A.;Motilva,M.-J。羟基酪醇和源自初榨橄榄油摄入的结肠代谢物诱导结肠癌细胞细胞周期停滞和凋亡。J. Agric. 食品化学。2017,65,6467–6476 。 [ Google Scholar ] [ CrossRef ]
- Sun, L.; Luo, C.; Liu, J. 羟基酪醇通过 ROS 生成诱导人类结肠癌细胞凋亡。食品功能 。2014 年,5,1909-1914年。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Terzuoli,E.;Giachetti,A. ; Ziche,M.;Donnini,S. 羟基酪醇是一种来自橄榄油的产品,它通过增强表皮生长因子受体降解来减缓结肠癌的生长。Mol . Nut. Food Res. 2016,60,519–529。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- Bernini,R.;Carastro,I.;Palmini,G.;Tanini,A.;Zonefrati,R.;Pinelli,P.;Brandi,ML;Romani,A. Olea europaea L. 中富含羟基酪醇的馏分的脂化。副产物及其对结直肠癌细胞模型的体外影响评估。J . Agric . Food Chem. 2017,65,6506–6512。[ Google Scholar ] [ CrossRef ]
- Rosignoli,P.;Fuccelli,R.;Sepporta,MV;Fabiani,R. 羟基酪醇的体外化学预防活性:特级初榨橄榄油中的主要酚类化合物。食品功能。2016 年,7,301-307。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Zubair,H.;Bhardwaj,A.;Ahmad,A.;Srivastava,SK;Khan,MA;Patel,GK;Singh,S.;Singh,AP 羟基酪醇诱导前列腺癌细胞凋亡和细胞周期停滞并抑制多种致癌信号通路。营养。癌症 2017,69,932-942 。 [ Google学术] [ CrossRef ]
- Calahorra,J.;Martínez-Lara,E.;De Dios , C.;Siles,E. 缺氧调节羟基酪醇在 MCF-7 乳腺癌细胞中的抗氧化作用。PLoS ONE 2018,13,e0203892。[ Google 学术搜索] [ CrossRef ]
- Zhao, B.; Ma, Y.; Xu, Z.; Wang, J.; Wang, F.; Wang, D.; Pan, S.; Wu, Y.; Pan, H.; Xu, D.; et al. 羟基酪醇是一种来自橄榄油的天然分子,它通过抑制 AKT 和核因子 κB 通路来抑制人类肝细胞癌细胞的生长。Cancer Lett. 2014 , 347 , 79–87。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- Tutino,V.;Caruso,MG;Messa,C.;Perri,E.;Notarnicola,M. 羟基酪醇对人类肝癌 HepG2 和 Hep3B 细胞系的抗增殖、抗氧化和抗炎作用。抗癌研究。2012 年,5371-5378。[ Google 学术]
- Li, S.; Han, Z.; Ma, Y.; Song, R.; Pei, T.; Zheng, T.; Wang, J.; Xu, D.; Fang, X.; Jiang, H.; et al. 羟基酪醇抑制胆管癌肿瘤生长:一项体内和体外研究。Oncol . Rep. 2014 , 31 , 145–152。[ Google 学术搜索] [ CrossRef ]
- Goldsmith,C.;Bond,D.;Jankowski,H.;Weidenhofer,J.;Stathopoulos,C.;Roach,P.;Scarlett,C. 橄榄生物酚橄榄苦苷和羟基酪醇选择性地减少增殖、影响细胞周期并诱导胰腺癌细胞凋亡。Int. J. Mol. Sci. 2018,19,1937 。 [ Google Scholar ] [ CrossRef ] [ PubMed ]
- 托特达,G.;卢皮纳奇,S.;维扎,D.;博诺菲利奥,R.;佩里,E.;博诺菲利奥,M.;洛法罗,D.;拉鲁萨,A.;莱昂,F.;吉利奥蒂,P.;等人。高剂量的羟基酪醇诱导乳头状和滤泡状甲状腺癌细胞凋亡。J.内分泌。投资。 2017 , 40 , 153–162。 [谷歌学术] [交叉引用] [ PubMed ]
- De las Hazas, MCL;Rubio, L.;Macia , A.;Motilva, MJ 羟基酪醇:潜在治疗应用的新兴趋势。Curr . Pharml. Desig 2018,24,2157–2179 。 [ Google 学术搜索] [ CrossRef ]
- Orsini,F.;Ami , D.;Lascialfari,A.;Natalello,A. 羟基酪醇和多巴胺对溶菌酶纤维形成的抑制:原子力显微镜研究。国际生物大分子杂志。2018,111,1100–1105 。[ Google学术搜索] [ CrossRef ]
- Goldstein, DS; Jinsmaa, Y.; Sullivan, P.; Holmes, C.; Kopin, IJ; Sharabi, Y. 3,4-二羟基苯乙醇(羟基酪醇)减轻 PC12 细胞中单胺氧化酶抑制过程中多巴胺自发氧化的增加。神经化学研究。2016,41,2173–2178 。 [ Google学术搜索] [ CrossRef ] [ PubMed ] [绿色版本]
- Hornedo-Ortega,R.;Cerezo,AB;Troncoso,AM;Garcia-Parrilla,MC 羟基酪醇对 PC12 细胞和原纤维形成的 α-突触核蛋白毒性的保护作用。食品化学毒理学。2018 年,120,41-49。[ Google 学术搜索] [ CrossRef ]
- 船小日-塔戈,M.;坂田,T.;藤原,S.;坂仓,A.;菅井,T.;塔戈,K.; Tamura, H. 羟基酪醇丁酸酯通过激活 SH-SY5Y 细胞中的 Nrf2/HO-1 轴来抑制 6-OHDA 诱导的细胞凋亡。欧元。 J.Pharmacol。 2017 , 834 , 246–256。 [谷歌学术] [交叉引用]
- Zheng, A.; Li, H.; Cao, K.; Xu, J.; Zou, X.; Li, Y.; Chen, C.; Liu, J.; Feng, Z. 母体补充羟基酪醇可改善产前应激后代的神经发生和认知功能。J . Nutr. Biochem. 2015 , 26 , 190–199。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- Davinelli, S.;Maes, M.;Corbi, G.;Zarrelli, A.;Willcox, DC;Scapagnini, G. 饮食中的植物化学物质和神经炎症:从机制洞察到转化挑战。Immun . Ageing 2016,13,16。[ Google Scholar ] [ CrossRef ]
- Priore,P.;Gnoni,A.;Natali,F.;Testini,M.;Gnoni,GV;Siculella,L.;Damiano,F. 油酸和羟基酪醇抑制 C6 胶质瘤细胞中的胆固醇和脂肪酸合成。氧化医学细胞长效。2017 年,2017 年,9076052。[ Google 学术搜索] [ CrossRef ]
- Carito, V.;Ceccanti, M.;Cestari, V.;Natella, F.;Bello, C.;Coccurello, R.;Mancinelli, R.;Fiore, M. 橄榄多酚对慢性乙醇成瘾小鼠模型的影响。营养学 2017,33,65–69。[ Google 学术搜索] [ CrossRef ] [绿色版本]
- Jemai, H.;El Feki, A.;Sayadi, S. 橄榄叶中的羟基酪醇和橄榄苦苷对链脲佐菌素糖尿病大鼠的抗糖尿病和抗氧化作用。J . Agric. 食品化学。2009 年,57,8798–8804。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- 安东尼奥·洛佩兹-维洛德雷斯,J.;阿卜杜勒·卡里姆,M.;佩德罗·德拉克鲁斯,J.;多洛雷斯·罗德里格斯-佩雷斯,M.;胡里奥·雷耶斯,J.;古兹曼-莫斯科索,R.;罗德里格斯-古铁雷斯,G.;费尔南德斯-博拉诺斯,J.; Antonio Gonzalez-Correa, J. 羟基酪醇对实验性糖尿病心血管生物标志物的影响。J.努特尔。生物化学。 2016 , 37 , 94–100。 [谷歌学术] [交叉引用] [绿色版]
- 雷耶斯,JJ;维拉纽瓦,B.;洛佩兹-维洛德雷斯,JA;德拉克鲁兹,太平绅士;罗梅罗,L.;罗德里格斯-佩雷斯,医学博士;罗德里格斯-古铁雷斯,G.;费尔南德斯-博拉诺斯,J.; Gonzalez-Correa,JA 羟基酪醇对实验性糖尿病的神经保护作用。J.阿格里克。食品化学。 2017 , 65 , 4378–4383。 [谷歌学术] [交叉引用]
- Suribabu,R.;Pindiprolu,SSS;Talluri,SV;Chintamaneni,P.;Samidala,N. 羟基酪醇对啮齿动物糖尿病周围神经病变的保护作用:抗氧化和抗炎作用的意义。拉丁美洲药学杂志。2017 年, 36,373–379 。[ Google 学术]
- Carmen Crespo,M.;Tome-Carneiro,J.;Pintado,C.;Davalos,A.;Visioli,F.;Burgos-Ramos,E. 羟基酪醇可在阿尔茨海默病星形胶质细胞模型中恢复正常的胰岛素信号传导。Biofactors 2017,43,540–548 。 [ Google学术搜索] [ CrossRef ]
- Soto-Alarcon, SA;Valenzuela, R.;Valenzuela, A.;Videla, LA 特级初榨橄榄油的保肝作用:其化学成分与参与保护的细胞信号通路之间的相互作用。内分泌代谢免疫紊乱药物靶点 2017,75–84。[ Google 学术搜索] [ CrossRef ]
- 皮罗齐,C.;喇嘛,A.;西梅奥利,R.;帕切洛,O.;结核病帕加诺;莫利卡,议员;迪吉达,F.;罗素,R.;马廖卡,S.;卡纳尼,RB;等人。羟基酪醇可预防 NAFLD 大鼠模型中的代谢损伤,从而减少肝脏炎症并恢复十二指肠完整性。J.努特尔。生物化学。 2016 , 30 , 108–115。 [谷歌学术] [交叉引用]
- Valenzuela, R.; Echeverria, F.; Ortiz, M.; Rincón-Cervera, M.Á.; Espinosa, A.; Hernandez-Rodas, MC; Illesca, P.; Valenzuela, A.; Videla, LA 羟基酪醇可防止高脂饮食喂养的小鼠不同组织中 Δ-5 和 Δ-6 去饱和酶肝脏活性降低、氧化应激和长链多不饱和脂肪酸含量消耗。Lipids Health Dis。2017 年,64–80。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Echeverría, F.; Valenzuela, R.; Bustamante, A.; Álvarez, D.; Ortiz, M.; Soto-Alarcon, SA; Muñoz, P.; Corbari, A.; Videla, LA 联合补充羟基酪醇-(HT-)二十碳五烯酸可减轻高脂饮食引起的大鼠肝脏氧化应激和脂肪变性,主要依赖于 HT。氧化。医学。细胞长寿。2018 年,2018 年,1-13。[ Google 学术] [ CrossRef ] [ PubMed ]
- Lemonakis,N.;Poudyal,H.;Halabalaki,M.;Brown,L.;Tsarbopoulos,A.;Skaltsounis,A.-L.;Gikas,E. 基于 LC-MS 的大鼠羟基酪醇给药代谢组学揭示了代谢综合征的改善。J . Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017,1041,45–59。[ Google 学术搜索] [ CrossRef ]
- Warnke, I.;Goralczyk, R.;Fuhrer, E.;Schwager, J. 饮食成分可减少小鼠 C3H10 T1/2 脂肪细胞中的脂质积累:一种量化脂肪滴的新型荧光方法。营养代谢。2011,8,30 。 [ Google学术搜索] [ CrossRef ]
- Dagla,I.;Benaki,D.;Baira,E.;Lemonakis,N.;Poudyal,H.;Brown,L.;Tsarbopoulos,A.;Skaltsounis,A.-L.;Mikros,E.;Gikas,E. 羟基酪醇治疗后代谢综合征大鼠肝脏代谢组的变化。基于质谱和核磁共振的代谢组学研究。Talanta 2018,178,246-257 。[ Google Scholar ] [ CrossRef ]
- Stefanon,B.;Colitti,M. 原始研究:羟基酪醇是橄榄油的一种成分,可减少甘油三酯的积累,并促进人类原代内脏脂肪细胞分化过程中的脂肪分解。Exp . Biol . Med. 2016,241,1796-1802。[ Google Scholar ] [ CrossRef ] [ PubMed ]
- 王娜;刘燕;马燕;温丹。羟基酪醇通过调节内质网应激改善胰岛素抵抗,并预防饮食诱导的肥胖小鼠的肝脂肪变性。J . Nutr. Biochem. 2018,57,180–188。[ Google Scholar ] [ CrossRef ]
- Illesca,P.;Valenzuela,R.;Espinosa,A.;Echeverría,F.;Soto-Alarcon,S.;Ortiz,M.;Videla,LA 羟基酪醇补充剂可通过恢复转录因子 Nrf2、SREBP-1c、PPAR-γ 和 NF-κB 改善高脂饮食小鼠白色脂肪组织的代谢紊乱。生物医学。药物治疗学 。2019,109,2472–2481。[ Google 学术搜索] [ CrossRef ]
- Valenzuela, R.; Illesca, P.; Echeverria, F.; Espinosa, A.; Rincon-Cervera, MA; Ortiz, M.; Hernandez-Rodas, MC; Valenzuela, A.; Videla, LA 羟基酪醇在小鼠肝脏高脂饮食诱发的致病改变中有益作用的分子适应性:PPAR-alpha 和 Nrf2 激活,以及 NF-kappa B 下调。食物功能。2017,8,1526–1537 。 [ Google学术搜索] [ CrossRef ]
- Bisignano,G.;Tomaino,A.;Lo Cascio,R.;Crisafi , G.;Uccella,N.;Saija,A. 橄榄苦苷和羟基酪醇的体外抗菌活性。J . Pharm. Pharmacol. 1999,51,971-974。[ Google 学术搜索] [ CrossRef ]
- Medina, E.;De Castro, A.;Romero , C.;Brenes, M. 橄榄油和其他植物油中酚类化合物浓度的比较:与抗菌活性的关系。J . Agric. Food Chem. 2006,54,4954–4961。[ Google Scholar ] [ CrossRef ]
- Wu, H.; Jiang, K.; Zhang, T.; Zhao, G.; Deng, G. 羟基酪醇通过抑制 Toll 样受体 2 和 TLR 2 下游通路在金黄色葡萄球菌诱发的小鼠乳腺炎中发挥抗炎作用。J . Funct. Food 2017,35,595–604。[ Google Scholar ] [ CrossRef ]
- Ghalandari, M.;Naghmachi, M.;Oliverio, M.;Nardi, M.;Shirazi, HRG;Eilami, O. 羟基酪醇、羟基酪醇乙酸酯和羟基酪醇油酸酯对金黄色葡萄球菌和表皮葡萄球菌的抗菌作用。Elect . J. Gen. Med. 2018,15,em46。[ Google 学术搜索] [ CrossRef ]
- Sogawa,K.;Kobayashi,M.;Suzuki,J.;Sanda,A.;Kodera,Y.;Fukuyama,M. 羟基酪醇对链球菌溶血素 O 诱导溶血的抑制活性。生物防治。科学 。2018年,23,77-80。[ Google 学术搜索] [ CrossRef ]
- Eilami,O.;Oliverio,M.;Hosseinian,S.;Motlagh,AH;Naghmachi,M. 橄榄叶提取物羟基酪醇对痤疮丙酸杆菌的抗菌作用。世界家庭医学杂志。2017 年,15,187-191。[ Google 学术搜索] [ CrossRef ]
- Furneri, PM;Piperno, A.;Sajia, A.;Bisignano, G. 羟基酪醇的抗支原体活性。抗微生物剂化疗。2004 年,48,4892–4894。[ Google 学术搜索] [ CrossRef ]
- Medina-Martinez,MS;Truchado,P. ;Castro -Ibanez,I.;Allende,A. 羟基酪醇的抗菌活性:当前的争议。生物科学。生物技术。生物化学。2016,80,801-810 。 [ Google 学术搜索] [ CrossRef ]
- 贝多亚,LM;贝尔特兰,M.;奥布雷贡-卡尔德隆,P.;加西亚-佩雷斯,J.;德拉托雷,HE;冈萨雷斯,N.;佩雷斯·奥尔梅达,M.;奥农,D.;卡帕,L.;戈麦斯-阿塞博,E.;等人。羟基酪醇:一类新型杀菌剂,具有广泛的抗 HIV-1 活性。2016 年艾滋病 ,30,2767–2776。 [谷歌学术] [交叉引用]
- Yamada, K.; Ogawa, H.; Hara, A.; Yoshida, Y.; Yonezawa, Y.; Karibe, K.; Nghia, VB; Yoshimura, H.; Yamamoto, Y.; Yamada, M.; 等。羟基酪醇对流感病毒的抗病毒作用机制似乎与病毒的形态变化有关。抗病毒研究 。2009,83,35–44。[ Google 学术搜索] [ CrossRef ]
- 迪亚利纳斯,G.;拉斐利杜,N.;卡尔帕克齐,I.;科米亚努,AC;楚瓦利,V.;赞察,I.;米克罗斯,E.;阿拉巴马州斯卡尔特索尼斯; Kostakis,IK 羟基酪醇 (HT) 类似物通过直接破坏真菌细胞膜发挥有效的抗真菌作用。正面。微生物。 2018 , 9 , 2624. [ Google Scholar ] [ CrossRef ] [ PubMed ] [绿色版]
- Belmonte-Reche,E.;Martinez-Garcia,M.;Penalver,P.;Gomez-Perez,V.;Lucas,R.;Gamarro,F.;Maria Perez-Victoria,J.;Carlos Morales,J. 酪醇和羟基酪醇衍生物作为抗锥虫和抗利什曼原虫药物。欧洲医学化学杂志。2016,119,132-140 。 [ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Fernandez-Pastor,I.;Martinez-Garcia,M.;Medina-O'Donnell,M.;Rivas,F.;Martinez,A.;Perez-Victoria,JM;Parra,A. 羟基酪醇的 omega-羟基烷基碳酸酯衍生物作为抗锥虫剂的半合成。J . Nat . Prod. 2018,81,2075–2082。[ Google Scholar ] [ CrossRef ]
- Yonezawa, Y.; Miyashita, T.; Nejishima, H.; Takeda, Y.; Imai, K.; Ogawa, H. 橄榄衍生的羟基酪醇对脂多糖诱导的 RAW264.7 细胞炎症的抗炎作用。J . Vet. Med. Sci. 2018 , 80 , 1801–1807。[ Google 学术搜索] [ CrossRef ] [绿色版本]
- Angeles Rosillo,M.;Sanchez-Hidalgo,M.;Castejon,ML;Montoya,T.;Gonzalez-Benjumea,A.;Fernandez-Bolanos,JG;Alarcon-de-la-Lastra,C. 特级初榨橄榄油酚羟基酪醇和羟基酪醇乙酸酯可下调人类滑膜细胞中与关节侵蚀有关的介质的产生。J . Funct . Food 2017,36,27–33。[ Google Scholar ] [ CrossRef ]
- Cetrullo,S.;D'Adamo,S.;Guidotti,S.;Borzi,RM;Flamigni,F。羟基酪醇通过 sirtuin 1 依赖和不依赖的机制诱导自噬,防止氧化应激下的软骨细胞死亡。生物化学。生物物理学。Acta - Gen。Subj。2016,1860,1181-1191 。[ Google 学术搜索] [ CrossRef ]
- Aparicio-Soto,M.;Sanchez-Hidalgo,M.;Cardeno,A.;Gonzalez-Benjumea,A.;Fernandez-Bolanos,JG;Alarcon-de-la-Lastra,C. 膳食补充羟基酪醇和羟基酪氨酸乙酸酯可预防小鼠由 pristane 诱发的系统性红斑狼疮。J . Funct. Food 2017,29,84-92 。 [ Google Scholar ] [ CrossRef ]
- 桑切斯-菲达尔戈,S.;维勒加斯,I.;阿帕里西奥·索托,M.;卡德诺,A.;马萨诸塞州罗西洛;冈萨雷斯-本朱梅亚,A.;马塞特,A.;洛佩斯,Ó。玛雅,I。费尔南德斯-博拉尼奥斯,JG;等人。膳食初榨橄榄油多酚:羟基酪氨酰乙酸酯和 3,4-二羟基苯基乙二醇对 DSS 诱导的小鼠急性结肠炎的影响。J.努特尔。生物化学。 2015 , 26 , 513–520。 [谷歌学术] [交叉引用]
- Yang, X.; Jing, T.; Li, Y.; He, Y.; Zhang, W.; Wang, B.; Zhang, J.; Wei, J.; Li, R. 羟基酪醇通过调节自噬和 Sirtuin 表达减轻小鼠 LPS 诱导的急性肺损伤。Curr . Mol. Med. 2017 , 17 , 149–159。[ Google 学术搜索] [ CrossRef ]
- Hagiwara, K.;Goto, T.;Araki, M.;Miyazaki, H.;Hagiwara, H. 橄榄多酚羟基酪醇可防止骨质流失。欧洲药理学杂志。2011 年,662,78–84。[ Google 学术搜索] [ CrossRef ]
- 加西亚-马丁内斯,O.;德卢纳-贝尔托斯,E.;拉莫斯-托雷西拉斯,J.;鲁伊斯,C.;米利亚,E.;洛伦佐,ML;希门尼斯,B.;桑切斯-奥尔蒂斯,A.; Rivas, A. 特级初榨橄榄油中的酚类化合物刺激人类成骨细胞增殖。《公共图书馆一号》 2016 年11月e0150045。 [谷歌学术] [交叉引用]
- Bonura, A.; Vlah, S.; Longo, A.; Bulati, M.; Melis, MR; Cibella, F.; Colombo, P. 羟基酪醇调节健康受试者 PBMCs 中 Par j 1 诱导的 IL-10 产生。免疫生物学 2016 , 221 , 1374–1377。[ Google 学术搜索] [ CrossRef ]
- 萨卢奇,S.;布拉蒂尼,S.;巴蒂斯特利,M.;布翁坦波,F.;卡诺尼科,B.;马尔泰利,AM;帕帕,S.; Falcieri,E. 酪醇可防止受辐射的角质形成细胞凋亡。J.德马托尔。科学。 2015 , 80 , 61–68。 [谷歌学术] [交叉引用]
- Avola,R.;Graziano,ACE;Pannuzzo,G. ; Bonina , F.;Cardile,V. 橄榄果实中的羟基酪醇可防止蓝光对人类角质形成细胞和成纤维细胞造成的损伤。J . Cell Physiol. 2019,234,9065–9076。[ Google 学术搜索] [ CrossRef ]
- Martinez-Lara,E.;Pena,A.;Calahorra,J. ; Canuelo,A.;Siles,E. 羟基酪醇降低氧化和亚硝化应激水平,并通过肾脏缺氧细胞中的 HIF-1 独立机制促进血管生成。食物功能。2016,7,540-548 。 [ Google 学术搜索] [ CrossRef ]
- Visioli,F.;Galli,C.;Bornet,F.;Mattei , A.;Patelli , R.;Galli,G.;Caruso,D. 橄榄油酚类物质在人体中的吸收与剂量有关。FEBS Lett. 2000,468,159–160。[ Google 学术搜索] [ CrossRef ]
- De las Hazas,MCL;Godinho-Pereira,J.;Macia,A.;Almeida,AF;Ventura,MR;Motilva,M.-J.;Santos,CJ 大脑对羟基酪醇及其主要循环代谢物的吸收:对神经元细胞的保护潜力。J . Funct. Food 2018,46,107–110 。 [ Google Scholar ] [ CrossRef ]
- Domínguez-Perles,R.;Auñón,D.;Ferreres,F.;Gil-Izquierdo,A. 口服正常和高剂量羟基酪醇、羟基酪醇乙酸酯和 DOPAC 后,Sprague-Dawley 大鼠血浆和尿液代谢物的性别差异。欧洲营养学杂志。2017 年,56,215-224。[ Google 学术搜索] [ CrossRef ]
- Bernini,R.;Montani,MSG;Merendino,N. ; Romani,A.;Velotti,F. 羟基酪醇衍生化合物:开发用于癌症预防和治疗的新药理学药剂的基础。J . Med. Chem. 2015,58,9089–9107。[ Google Scholar ] [ CrossRef ]
- Mateos,R.;Pereira-Caro,G.;Saha,S.;Cert,R.;Redondo-Horcajo,M.;Bravo,L.;Kroon,PA 羟基酪醇的乙酰化增强了其在分化的 Caco-2 细胞单层中的运输。食品化学。2011,125,865-872 。 [ Google 学术搜索] [ CrossRef ]
- Pereira-Caro,G.;Mateos,R.;Shikha,S.;Andres,M.;José Luis,E.;Laura,B.;Paul,AK 肠细胞样 Caco-2/TC7 细胞对羟基酪醇新型亲脂性醚衍生物的跨上皮运输和代谢。J . Agric . 食品化学。2010,58,11501–11509 。 [ Google 学术搜索] [ CrossRef ]
- Pereira-Caro,G.;Madrona,A.;Bravo,L.;Espartero,JL;Alcudia,F.;Cert,A.;Mateos,R. 烷基羟基酪氨酸醚(一类新型羟基酪醇衍生物)的抗氧化活性评价。食品化学。2009 年,115,86-91。[ Google 学术搜索] [ CrossRef ]
- Grasso,S.;Siracusa,L.;Spatafora, C .; Renis ,M.;Tringali,C. 羟基酪醇亲脂性类似物:酶合成、自由基清除活性和 DNA 氧化损伤保护。Bioorg . Chem. 2007,35,137-152。[ Google 学术搜索] [ CrossRef ] [ PubMed ]
- Manna, C.; Galletti, P.; Maisto, G.; Cucciolla, V.; D'Angelo, S.; Zappia, V. 橄榄油羟基酪醇在 Caco-2 细胞中的运输机制和代谢。FEBS Lett. 2000 , 470 , 341–344。[ Google Scholar ] [ CrossRef ] [绿色版本]
- 牧师,A.;罗德里格斯·莫拉托,J.;奥莱斯蒂,E.;普哈达斯,M.;佩雷斯·马纳 (Pérez-Mañá),C.;希门涅茨,O.;菲托,M.;科瓦斯,M.-I.;太阳的。;莫蒂尔瓦,M.-J.;等人。施用橄榄油后人血浆中游离羟基酪醇的分析。J. Chromatogr。 2016 , 1437 , 183–190。 [谷歌学术] [交叉引用] [绿色版]
- Tuck, KL;Hayball, PJ;Stupans, I. 大鼠体内橄榄油中主要酚类成分羟基酪醇代谢物的结构表征。J . Agric. 食品化学。2002 年,50,2404–2409。[ Google 学术搜索] [ CrossRef ]
- Visioli,F.;Galli,C. ; Grande,S.;Colonnelli,K.;Patelli ,C.;Galli,G.;Caruso,D. 羟基酪醇排泄在大鼠和人类之间有所不同,并且取决于给药载体。J . Nutr。2003,133,2612-2615 。 [ Google Scholar ] [ CrossRef ]
- González-Santiago, M.; Fonollá, J.; Lopez-Huertas, E. Human absorption of a supplement containing purified hydroxytyrosol, a natural antioxidant from olive oil, and evidence for its transient association with low-density lipoproteins. Pharmacoll. Res. 2010, 61, 364–370. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, S.; Manna, C.; Migliardi, V.; Mazzoni, O.; Morrica, P.; Capasso, G.; Pontoni, G.; Galletti, P.; Zappia, V. Pharmacokinetics and metabolism of hydroxytyrosol, a natural antioxidant from olive oil. Drug Metab. Dispos. 2001, 29, 1492–1498. [Google Scholar] [PubMed]
- Fernandez-Avila, C.; Montes, R.; Castellote, A.I.; Chisaguano, A.M.; Fito, M.; Covas, M.I.; Munoz-Aguallo, D.; Nyyssonen, K.; Zunft, H.J.; Lopez-Sabater, M.C. Fast determination of virgin olive oil phenolic metabolites in human high-density lipoproteins. Biomed. Chromatogr. 2015, 29, 1035–1041. [Google Scholar] [CrossRef]
- Serra, A.; Rubió, L.; Borràs, X.; Macià, A.; Romero, M.-P.; Motilva, M.-J. Distribution of olive oil phenolic compounds in rat tissues after administration of a phenolic extract from olive cake. Mol. Nutr. Food Res. 2012, 56, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.; Asseburg, H.; Kuntz, S.; Muller, W.E.; Eckert, G.P. Effects of Polyphenols on Brain Ageing and Alzheimer’s Disease: Focus on Mitochondria. Mol. Neurobiol. 2012, 46, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.; Müller, W.E.; Eckert, G.P. Cytoprotective effects of olive mill wastewater extract and its main constituent hydroxytyrosol in PC12 cells. Pharmacol. Res. 2010, 62, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morató, J.; Boronat, A.; Kotronoulas, A.; Pujadas, M.; Pastor, A.; Olesti, E.; Pérez-Mañá, C.; Khymenets, O.; Fitó, M.; Farré, M.; et al. Metabolic disposition and biological significance of simple phenols of dietary origin: Hydroxytyrosol and tyrosol. Drug Metabol. Rev. 2016, 48, 218–236. [Google Scholar] [CrossRef]
- De la Torre, R.; Covas, M.I.; Pujadas, M.A.; Fitó, M.; Farré, M. Is dopamine behind the health benefits of red wine? Eur. J. Nutr. 2006, 45, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.L.; Sim, M.K. Reduction of dihydroxyphenylacetic acid by a novel enzyme in the rat brain. Biochem. Pharmacol. 1995, 50, 1333–1337. [Google Scholar] [CrossRef]
- Perez-Mana, C.; Farre, M.; Pujadas, M.; Mustata, C.; Menoyo, E.; Pastor, A.; Langohr, K.; de la Torre, R. Ethanol induces hydroxytyrosol formation in humans. Pharmacol. Res. 2015, 95–96, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Davis, V.E.; Walsh, M.J. Alcohol, amines, and alkaloids: A possible biochemical basis for alcohol addiction. Science 1970, 167, 1005–1007. [Google Scholar]
- Davis, V.E.; Walsh, M.J.; Yamanaka, Y. Augmentation of alkaloid formation from dopamine by alcohol and acetaldehyde in vitro. J. Pharmacol. Exp. Ther. 1970, 174, 401–412. [Google Scholar] [PubMed]
- Tank, A.W.; Weiner, H. Ethanol-induced alteration of dopamine metabolism in rat liver. Biochem. Pharmacol. 1979, 28, 3139–3147. [Google Scholar] [CrossRef]
- Marchitti, S.A.; Deitrich, R.A.; Vasiliou, V. Neurotoxicity and Metabolism of the Catecholamine-Derived 3,4-Dihydroxyphenylacetaldehyde and 3,4-Dihydroxyphenylglycolaldehyde: The Role of Aldehyde Dehydrogenase. Pharmacol. Rev. 2007, 59, 125–150. [Google Scholar] [CrossRef]
- Rubió, L.; Macià, A.; Valls, R.M.; Pedret, A.; Romero, M.-P.; Solà, R.; Motilva, M.-J. A new hydroxytyrosol metabolite identified in human plasma: Hydroxytyrosol acetate sulphate. Food Chem. 2012, 134, 1132–1136. [Google Scholar] [CrossRef]
- De las Hazas, M.C.L.; Rubió, L.; Kotronoulas, A.; de la Torre, R.; Solà, R.; Motilva, M.-J. Dose effect on the uptake and accumulation of hydroxytyrosol and its metabolites in target tissues in rats. Mol. Nutr. Food Res. 2015, 59, 1395–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De las Hazas, M.C.L.; Piñol, C.; Macià, A.; Romero, M.-P.; Pedret, A.; Solà, R.; Rubió, L.; Motilva, M.-J. Differential absorption and metabolism of hydroxytyrosol and its precursors oleuropein and secoiridoids. J. Funct. Food 2016, 22, 52–63. [Google Scholar] [CrossRef] [Green Version]
- De las Hazas, M.C.L.; Motilva, M.J.; Piñol, C.; Macià, A. Application of dried blood spot cards to determine olive oil phenols (hydroxytyrosol metabolites) in human blood. Talanta 2016, 159, 189–193. [Google Scholar] [CrossRef]
- Caruso, D.; Visioli, F.; Patelli, R.; Galli, C.; Galli, G. Urinary excretion of olive oil phenols and their metabolites in humans. Metabolism 2001, 50, 1426–1428. [Google Scholar] [CrossRef]
- Kotronoulas, A.; Pizarro, N.; Serra, A.; Robledo, P.; Joglar, J.; Rubió, L.; Hernaéz, Á.; Tormos, C.; Motilva, M.J.; Fitó, M.; et al. Dose-dependent metabolic disposition of hydroxytyrosol and formation of mercapturates in rats. Pharmacol. Res. 2013, 77, 47–56. [Google Scholar] [CrossRef]
- Wu, Y.-T.; Lin, L.-C.; Tsai, T.-H. Measurement of free hydroxytyrosol in microdialysates from blood and brain of anesthetized rats by liquid chromatography with fluorescence detection. J. Chromatogr. A 2009, 1216, 3501–3507. [Google Scholar] [CrossRef]
- Mateos, R.; Martínez-López, S.; Baeza Arévalo, G.; Amigo-Benavent, M.; Sarriá, B.; Bravo-Clemente, L. Hydroxytyrosol in functional hydroxytyrosol-enriched biscuits is highly bioavailable and decreases oxidised low density lipoprotein levels in humans. Food Chem. 2016, 205, 248–256. [Google Scholar] [CrossRef]
- Serreli, G.; Deiana, M. Biological Relevance of Extra Virgin Olive Oil Polyphenols Metabolites. Antioxidants 2018, 7, 170. [Google Scholar] [CrossRef]
- García-Villalba, R.; Larrosa, M.; Possemiers, S.; Tomás-Barberán, F.A.; Espín, J.C. Bioavailability of phenolics from an oleuropein-rich olive (Olea europaea) leaf extract and its acute effect on plasma antioxidant status: Comparison between pre- and postmenopausal women. Eur. J. Nutr. 2014, 53, 1015–1027. [Google Scholar] [CrossRef]
- Mosele, J.I.; Martín-Peláez, S.; Macià, A.; Farràs, M.; Valls, R.-M.; Catalán, Ú.; Motilva, M.-J. Faecal microbial metabolism of olive oil phenolic compounds: In vitro and in vivo approaches. Mol. Nutr. Food Res. 2014, 58, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.; Macià, A.; Motilva, M.-J. Metabolic and Microbial Modulation of the Large Intestine Ecosystem by Non-Absorbed Diet Phenolic Compounds: A Review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [CrossRef]
- Suárez, M.; Valls, R.M.; Romero, M.-P.; Macià, A.; Fernández, S.; Giralt, M.; Solà, R.; Motilva, M.-J. Bioavailability of phenols from a phenol-enriched olive oil. Br. J. Nutr. 2011, 106, 1691–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chashmi, N.A.; Emadi, S.; Khastar, H. Protective effects of hydroxytyrosol on gentamicin induced nephrotoxicity in mice. Biochem. Biophys. Res. Commun. 2017, 482, 1427–1429. [Google Scholar] [CrossRef]
- Miró-Casas, E.; Farré Albaladejo, M.; Covas, M.-I.; Rodriguez, J.O.; Menoyo Colomer, E.; Lamuela Raventós, R.M.; de la Torre, R. Capillary Gas Chromatography–Mass Spectrometry Quantitative Determination of Hydroxytyrosol and Tyrosol in Human Urine after Olive Oil Intake. Anal. Biochem. 2001, 294, 63–72. [Google Scholar] [CrossRef]
- Miró-Casas, E.; Covas, M.-I.; Fitó, M.; Farré-Albadalejo, M.; Marrugat, J.; de la Torre, R. Tyrosol and hydroxytyrosol are absorbed from moderate and sustained doses of virgin olive oil in humans. Eur. J. Clin. Nutr. 2003, 57, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Khymenets, O.; Fito, M.; Tourino, S.; Munoz-Aguayo, D.; Pujadas, M.; Torres, J.L.; Joglar, J.; Farre, M.; Covas, M.-I.; de la Torre, R. Antioxidant Activities of Hydroxytyrosol Main Metabolites Do Not Contribute to Beneficial Health Effects after Olive Oil Ingestion. Drug Metab. Dispos. 2010, 38, 1417–1421. [Google Scholar] [CrossRef] [Green Version]
- Giordano, E.; Dangles, O.; Rakotomanomana, N.; Baracchini, S.; Visioli, F. 3-O-Hydroxytyrosol glucuronide and 4-O-hydroxytyrosol glucuronide reduce endoplasmic reticulum stress in vitro. Food Funct. 2015, 6, 3275–3281. [Google Scholar] [CrossRef]
- Atzeri, A.; Lucas, R.; Incani, A.; Peñalver, P.; Zafra-Gómez, A.; Melis, M.P.; Pizzala, R.; Morales, J.C.; Deiana, M. Hydroxytyrosol and tyrosol sulfate metabolites protect against the oxidized cholesterol pro-oxidant effect in Caco-2 human enterocyte-like cells. Food Funct. 2016, 7, 337–346. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Silva, A.; Almeida, V.; Carvalheira, M.; Serra, C.; Rodrígues-Borges, J.E.; Fernandes, J.; Belo, L.; Santos-Silva, A. Protective Activity of Hydroxytyrosol Metabolites on Erythrocyte Oxidative-Induced Hemolysis. J. Agric. Food Chem. 2013, 61, 6636–6642. [Google Scholar] [CrossRef]
- Deiana, M.; Incani, A.; Rosa, A.; Atzeri, A.; Loru, D.; Cabboi, B.; Paola Melis, M.; Lucas, R.; Morales, J.C.; Assunta Dessì, M. Hydroxytyrosol glucuronides protect renal tubular epithelial cells against H2O2 induced oxidative damage. Chem. Biol. Interact. 2011, 193, 232–239. [Google Scholar] [CrossRef]
- Di Benedetto, R.; Varì, R.; Scazzocchio, B.; Filesi, C.; Santangelo, C.; Giovannini, C.; Matarrese, P.; D’Archivio, M.; Masella, R. Tyrosol, the major extra virgin olive oil compound, restored intracellular antioxidant defences in spite of its weak antioxidative effectiveness. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 535–545. [Google Scholar] [CrossRef]
- Lee, K.M.; Hur, J.; Lee, Y.; Yoon, B.-R.; Choi, S.Y. Protective Effects of Tyrosol Against Oxidative Damage in L6 Muscle Cells. Food Sci. Technol. Res. 2018, 24, 943–947. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-Y.; Huang, I.-T.; Shih, H.-J.; Chang, Y.-Y.; Kao, M.-C.; Shih, P.-C.; Huang, C.-J. Cluster of differentiation 14 and toll-like receptor 4 are involved in the anti-inflammatory effects of tyrosol. J. Funct. Food 2019, 53, 93–104. [Google Scholar] [CrossRef]
- Samuel, S.M.; Thirunavukkarasu, M.; Penumathsa, S.V.; Paul, D.; Maulik, N. Akt/FOXO3a/SIRT1-Mediated Cardioprotection by n-Tyrosol against Ischemic Stress in Rat in Vivo Model of Myocardial Infarction: Switching Gears toward Survival and Longevity. J. Agric. Food Chem. 2008, 56, 9692–9698. [Google Scholar] [CrossRef]
- Plotnikov, M.B.; Aliev, O.; Sidekhmenova, A.; Shamanaev, A.Y.; Anishchenko, A.M.; Fomina, T.; Plotnikova, T.M.; Arkhipov, A.M. Effect of p-tyrosol on hemorheological parameters and cerebral capillary network in young spontaneously hypertensive rats. Microvasc. Res. 2018, 119, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Osipenko, A.N.; Plotnikova, T.M.; Chernysheva, G.A.; Smolyakova, V.I. The mechanisms of neuroprotective action of p-tyrosol after the global cerebral ischemia in rats. Byulleten Sibirskoy Meditsiny 2017, 16, 65–72. [Google Scholar] [CrossRef]
- Chandramohan, R.; Pari, L. Anti-inflammatory effects of tyrosol in streptozotocin-induced diabetic Wistar rats. J. Funct. Food 2016, 27, 17–28. [Google Scholar] [CrossRef]
- Lee, H.; Im, S.W.; Jung, C.H.; Jang, Y.J.; Ha, T.Y.; Ahn, J. Tyrosol, an olive oil polyphenol, inhibits ER stress-induced apoptosis in pancreatic beta-cell through JNK signaling. Biochem. Biophys. Res. Commun. 2016, 469, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Priore, P.; Siculella, L.; Gnoni, G.V. Extra virgin olive oil phenols down-regulate lipid synthesis in primary-cultured rat-hepatocytes. J. Nutr. Biochem. 2014, 25, 683–691. [Google Scholar] [CrossRef]
- Sarna, L.K.; Sid, V.; Wang, P.; Siow, Y.L.; House, J.D.; Karmin, O. Tyrosol Attenuates High Fat Diet-Induced Hepatic Oxidative Stress: Potential Involvement of Cystathionine β-Synthase and Cystathionine γ-Lyase. Lipids 2016, 51, 583–590. [Google Scholar] [CrossRef] [PubMed]
- St-Laurent-Thibault, C.; Arseneault, M.; Longpré, F.; Ramassamy, C. Tyrosol and hydroxytyrosol, two main components of olive oil, protect N2a cells against amyloid-β-induced toxicity. Involvement of the NF-κB signaling. Curr. Alzheimer Res. 2011, 8, 543–551. [Google Scholar] [CrossRef]
- Amini, A.; Liu, M.; Ahmad, Z. Understanding the link between antimicrobial properties of dietary olive phenolics and bacterial ATP synthase. Int. J. Biol. Macromol. 2017, 101, 153–164. [Google Scholar] [CrossRef]
- Puel, C.; Mardon, J.; Agalias, A.; Davicco, M.-J.; Lebecque, P.; Mazur, A.; Horcajada, M.-N.; Skaltsounis, A.-L.; Coxam, V. Major phenolic compounds in olive oil modulate bone loss in an ovariectomy/inflammation experimental model. J. Agric. Food Chem. 2008, 56, 9417–9422. [Google Scholar] [CrossRef]
- Cañuelo, A.; Gilbert-López, B.; Pacheco-Liñán, P.; Martínez-Lara, E.; Siles, E.; Miranda-Vizuete, A. Tyrosol, a main phenol present in extra virgin olive oil, increases lifespan and stress resistance in Caenorhabditis elegans. Mech Ageing Dev. 2012, 133, 563–574. [Google Scholar] [CrossRef]
- Lee, D.-H.; Kim, Y.-J.; Kim, M.; Ahn, J.; Ha, T.-Y.; Lee, S.; Jang, Y.; Jung, C. Pharmacokinetics of Tyrosol Metabolites in Rats. Molecules 2016, 21, 128. [Google Scholar] [CrossRef]
- Tacker, M.; Creaven, P.J.; McIsaac, W.M. Alteration in tyramine metabolism by ethanol. Biochem. Pharmacol. 1970, 19, 604–607. [Google Scholar] [CrossRef]
- Perez-Mana, C.; Farré, M.; Rodríguez-Morató, J.; Papaseit, E.; Pujadas, M.; Fitó, M.; Robledo, P.; Covas, M.-I.; Cheynier, V.; Meudec, E.; et al. Moderate consumption of wine, through both its phenolic compounds and alcohol content, promotes hydroxytyrosol endogenous generation in humans. A randomized controlled trial. Mol. Nutr. Food Res. 2015, 59, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morató, J.; Robledo, P.; Tanner, J.-A.; Boronat, A.; Pérez-Mañá, C.; Oliver Chen, C.-Y.; Tyndale, R.F.; de la Torre, R. CYP2D6 and CYP2A6 biotransform dietary tyrosol into hydroxytyrosol. Food Chem. 2017, 217, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Mateos, R.; Goya, L.; Bravo, L. Metabolism of the Olive Oil Phenols Hydroxytyrosol, Tyrosol, and Hydroxytyrosyl Acetate by Human Hepatoma HepG2 Cells. J. Agric. Food Chem. 2005, 53, 9897–9905. [Google Scholar] [CrossRef] [Green Version]
- Muriana, F.J.G.; Montserrat-de la Paz, S.; Lucas, R.; Bermudez, B.; Jaramillo, S.; Morales, J.C.; Abia, R.; Lopez, S. Tyrosol and its metabolites as antioxidative and anti-inflammatory molecules in human endothelial cells. Food Funct. 2017, 8, 2905–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassen, I.; Casabianca, H.; Hosni, K. Biological activities of the natural antioxidant oleuropein: Exceeding the expectation – A mini-review. J. Funct. Food 2015, 18, 926–940. [Google Scholar] [CrossRef]
- Visioli, F.; Bellomo, G.; Galli, C. Free Radical-Scavenging Properties of Olive Oil Polyphenols. Biochem. Biophys. Res. Commun. 1998, 247, 60–64. [Google Scholar] [CrossRef]
- Fabiani, R.; Rosignoli, P.; De Bartolomeo, A.; Fuccelli, R.; Servili, M.; Montedoro, G.F.; Morozzi, G. Oxidative DNA Damage Is Prevented by Extracts of Olive Oil, Hydroxytyrosol, and Other Olive Phenolic Compounds in Human Blood Mononuclear Cells and HL60 Cells. J. Nutr. 2008, 138, 1411–1416. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.A.; Zoubouli, P.; Calder, P.C. Differential anti-inflammatory effects of phenolic compounds from extra virgin olive oil identified in human whole blood cultures. Nutrition 2005, 21, 389–394. [Google Scholar] [CrossRef]
- Dell’Agli, M.; Fagnani, R.; Galli, G.V.; Maschi, O.; Gilardi, F.; Bellosta, S.; Crestani, M.; Bosisio, E.; De Fabiani, E.; Caruso, D. Olive Oil Phenols Modulate the Expression of Metalloproteinase 9 in THP-1 Cells by Acting on Nuclear Factor-κB Signaling. J. Agric. Food Chem. 2010, 58, 2246–2252. [Google Scholar] [CrossRef]
- Giner, E.; Andújar, I.; Recio, M.C.; Ríos, J.L.; Cerdá-Nicolás, J.M.; Giner, R.M. Oleuropein Ameliorates Acute Colitis in Mice. J. Agric. Food Chem. 2011, 59, 12882–12892. [Google Scholar] [CrossRef]
- Giner, E.; Recio, M.-C.; Ríos, J.-L.; Giner, R.-M. Oleuropein Protects against Dextran Sodium Sulfate-Induced Chronic Colitis in Mice. J. Nat. Prod. 2013, 76, 1113–1120. [Google Scholar] [CrossRef]
- Larussa, T.; Oliverio, M.; Suraci, E.; Greco, M.; Placida, R.; Gervasi, S.; Marasco, R.; Imeneo, M.; Paolino, D.; Tucci, L.; et al. Oleuropein Decreases Cyclooxygenase-2 and Interleukin-17 Expression and Attenuates Inflammatory Damage in Colonic Samples from Ulcerative Colitis Patients. Nutrients 2017, 9, 391. [Google Scholar] [CrossRef]
- Domitrović, R.; Jakovac, H.; Marchesi, V.V.; Šain, I.; Romić, Ž.; Rahelić, D. Preventive and therapeutic effects of oleuropein against carbon tetrachloride-induced liver damage in mice. Pharmacol. Res. 2012, 65, 451–464. [Google Scholar] [CrossRef]
- Shi, C.; Chen, X.; Liu, Z.; Meng, R.; Zhao, X.; Liu, Z.; Guo, N. Oleuropein protects L-02 cells against H2O2-induced oxidative stress by increasing SOD1, GPx1 and CAT expression. Biomed. Pharmacother. 2017, 85, 740–748. [Google Scholar] [CrossRef]
- Potočnjak, I.; Škoda, M.; Pernjak-Pugel, E.; Peršić, M.P.; Domitrović, R. Oral administration of oleuropein attenuates cisplatin-induced acute renal injury in mice through inhibition of ERK signaling. Mol. Nutr. Food Res. 2016, 60, 530–541. [Google Scholar] [CrossRef]
- Maalej, A.; Forte, M.; Bouallagui, Z.; Donato, S.; Mita, L.; Mita, D.G.; Filosa, S.; Crispi, S.; Sayadi, S. Olive compounds attenuate oxidative damage induced in HEK-293 cells via MAPK signaling pathway. J. Funct. Food 2017, 39, 18–27. [Google Scholar] [CrossRef]
- Castejón, M.L.; Rosillo, M.Á.; Montoya, T.; González-Benjumea, A.; Fernández-Bolaños, J.M.; Alarcón-de-la-Lastra, C. Oleuropein down-regulated IL-1β-induced inflammation and oxidative stress in human synovial fibroblast cell line SW982. Food Funct. 2017, 8, 1890–1898. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Choi, Y.-J.; Kang, M.-K.; Lee, E.-J.; Kim, D.Y.; Oh, H.; Kang, Y.-H. Oleuropein Curtails Pulmonary Inflammation and Tissue Destruction in Models of Experimental Asthma and Emphysema. J. Agric. Food Chem. 2018, 66, 7643–7654. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.M.; Arnal, C.; Navarro, M.A.; Osada, J. Efficacy of bioactive compounds from extra virgin olive oil to modulate atherosclerosis development. Mol. Nutr. Food Res. 2012, 56, 1043–1057. [Google Scholar] [CrossRef]
- Carluccio, M.A.; Siculella, L.; Ancora, M.A.; Massaro, M.; Scoditti, E.; Storelli, C.; Visioli, F.; Distante, A.; De Caterina, R. Olive Oil and Red Wine Antioxidant Polyphenols Inhibit Endothelial Activation: Antiatherogenic Properties of Mediterranean Diet Phytochemicals. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 622–629. [Google Scholar] [CrossRef]
- Dell’Agli, M.; Fagnani, R.; Mitro, N.; Scurati, S.; Masciadri, M.; Mussoni, L.; Galli, G.V.; Bosisio, E.; Crestani, M.; De Fabiani, E.; et al. Minor Components of Olive Oil Modulate Proatherogenic Adhesion Molecules Involved in Endothelial Activation. J. Agric. Food Chem. 2006, 54, 3259–3264. [Google Scholar] [CrossRef]
- Lockyer, S.; Corona, G.; Yaqoob, P.; Spencer, J.P.E.; Rowland, I. Secoiridoids delivered as olive leaf extract induce acute improvements in human vascular function and reduction of an inflammatory cytokine: A randomised, double-blind, placebo-controlled, cross-over trial. Br. J. Nutr. 2015, 114, 75–83. [Google Scholar] [CrossRef] [PubMed]
- De Bock, M.; Derraik, J.G.B.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) Leaf Polyphenols Improve Insulin Sensitivity in Middle-Aged Overweight Men: A Randomized, Placebo-Controlled, Crossover Trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef]
- Lockyer, S.; Rowland, I.; Spencer, J.P.E.; Yaqoob, P.; Stonehouse, W. Impact of phenolic-rich olive leaf extract on blood pressure, plasma lipids and inflammatory markers: A randomised controlled trial. Eur. J. Nutr. 2017, 56, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Susalit, E.; Agus, N.; Effendi, I.; Tjandrawinata, R.R.; Nofiarny, D.; Perrinjaquet-Moccetti, T.; Verbruggen, M. Olive (Olea europaea) leaf extract effective in patients with stage-1 hypertension: Comparison with Captopril. Phytomedicine 2011, 18, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Dell’Agli, M.; Maschi, O.; Galli, G.V.; Fagnani, R.; Dal Cero, E.; Caruso, D.; Bosisio, E. Inhibition of platelet aggregation by olive oil phenols via cAMP-phosphodiesterase. Br. J. Nutr. 2008, 99, 945–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, C.; Migliardi, V.; Golino, P.; Scognamiglio, A.; Galletti, P.; Chiariello, M.; Zappia, V. Oleuropein prevents oxidative myocardial injury induced by ischemia and reperfusion. J. Nutr. Biochem. 2004, 15, 461–466. [Google Scholar] [CrossRef]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Constantinou, M.; Agalias, A.; Magiatis, P.; Skaltsounis, A.L.; Kamber, E.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. The Olive Constituent Oleuropein Exhibits Anti-Ischemic, Antioxidative, and Hypolipidemic Effects in Anesthetized Rabbits. J. Nutr. 2006, 136, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Bai, Y.; Li, C.; Yang, K.; Wei, W.; Li, Z.; Pan, L.; Li, X.; Zhang, X. Oleuropein Protects Cardiomyocyte against Apoptosis via Activating the Reperfusion Injury Salvage Kinase Pathway In Vitro. J. Evid.-Based Complement. Altern. Med. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Jin, H.-X.; Zhang, Y.-H.; Guo, R.-N.; Zhao, S.-N. Inhibition of MEK/ERK/STAT3 signaling in oleuropein treatment inhibits myocardial ischemia/reperfusion. Int. J. Mol. Med. 2018, 42, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Andreadou, I.; Benaki, D.; Efentakis, P.; Bibli, S.-I.; Milioni, A.-I.; Papachristodoulou, A.; Zoga, A.; Skaltsounis, A.-L.; Mikros, E.; Iliodromitis, E. The Natural Olive Constituent Oleuropein Induces Nutritional Cardioprotection in Normal and Cholesterol-Fed Rabbits: Comparison with Preconditioning. Planta Med. 2015, 81, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Y.; Yang, Z.; Fang, K.; Shi, Z.-L.; Ren, D.-H.; Sun, J. Oleuropein prevents the development of experimental autoimmune myocarditis in rats. Internat. Immunopharmacol. 2017, 48, 187–195. [Google Scholar] [CrossRef]
- Shamshoum, H.; Vlavcheski, F.; Tsiani, E. Anticancer effects of oleuropein. BioFactors 2017, 43, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ahn, K.S.; Shanmugam, M.K.; Wang, H.; Shen, H.; Arfuso, F.; Chinnathambi, A.; Alharbi, S.A.; Chang, Y.; Sethi, G.; et al. Oleuropein induces apoptosis via abrogating NF-κB activation cascade in estrogen receptor-negative breast cancer cells. J. Cell Biochem. 2019, 120, 4504–4513. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.K.; Elamin, M.H.; Daghestani, M.H.; Omer, S.A.; Al-Olayan, E.M.; Elobeid, M.A.; Virk, P.; Mohammed, O.B. Oleuropein Induces Anti-metastatic Effects in Breast Cancer. Asian Pac. J. Cancer Prev. 2012, 13, 4555–4559. [Google Scholar] [CrossRef] [Green Version]
- Sepporta, M.V.; Fuccelli, R.; Rosignoli, P.; Ricci, G.; Servili, M.; Morozzi, G.; Fabiani, R. Oleuropein inhibits tumour growth and metastases dissemination in ovariectomised nude mice with MCF-7 human breast tumour xenografts. J. Funct. Food 2014, 8, 269–273. [Google Scholar] [CrossRef]
- Sherif, I.O.; Al-Gayyar, M.M.H. Oleuropein potentiates anti-tumor activity of cisplatin against HepG2 through affecting proNGF/NGF balance. Life Sci. 2018, 198, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Bakir, M.; Geyikoglu, F.; Koc, K.; Cerig, S. Therapeutic effects of oleuropein on cisplatin-induced pancreas injury in rats. J. Cancer Res. Ther. 2018, 14, 671–678. [Google Scholar]
- Ruzzolini, J.; Peppicelli, S.; Andreucci, E.; Bianchini, F.; Scardigli, A.; Romani, A.; la Marca, G.; Nediani, C.; Calorini, L. Oleuropein, the Main Polyphenol of Olea europaea Leaf Extract, Has an Anti-Cancer Effect on Human BRAF Melanoma Cells and Potentiates the Cytotoxicity of Current Chemotherapies. Nutrients 2018, 10, 1950. [Google Scholar] [CrossRef]
- Al-Azzawie, H.F.; Alhamdani, M.-S.S. Hypoglycemic and antioxidant effect of oleuropein in alloxan-diabetic rabbits. Life Sci. 2006, 78, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Murotomi, K.; Umeno, A.; Yasunaga, M.; Shichiri, M.; Ishida, N.; Koike, T.; Matsuo, T.; Abe, H.; Yoshida, Y.; Nakajima, Y. Oleuropein-Rich Diet Attenuates Hyperglycemia and Impaired Glucose Tolerance in Type 2 Diabetes Model Mouse. J. Agric. Food Chem. 2015, 63, 6715–6722. [Google Scholar] [CrossRef]
- Alkhateeb, H.; Al-Duais, M.; Qnais, E. Beneficial effects of oleuropein on glucose uptake and on parameters relevant to the normal homeostatic mechanisms of glucose regulation in rat skeletal muscle. Phytother. Res. 2018, 32, 651–656. [Google Scholar] [CrossRef]
- Wu, L.; Velander, P.; Liu, D.; Xu, B. Olive Component Oleuropein Promotes β-Cell Insulin Secretion and Protects β-Cells from Amylin Amyloid-Induced Cytotoxicity. Biochemistry 2017, 56, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-N.; Jung, J.-H.; Park, H.; Kim, H. Olive leaf extract suppresses messenger RNA expression of proinflammatory cytokines and enhances insulin receptor substrate 1 expression in the rats with streptozotocin and high-fat diet–induced diabetes. Nutr. Res. 2014, 34, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Wainstein, J.; Ganz, T.; Boaz, M.; Bar Dayan, Y.; Dolev, E.; Kerem, Z.; Madar, Z. Olive Leaf Extract as a Hypoglycemic Agent in Both Human Diabetic Subjects and in Rats. J. Med. Food 2012, 15, 605–610. [Google Scholar] [CrossRef]
- Carnevale, R.; Silvestri, R.; Loffredo, L.; Novo, M.; Cammisotto, V.; Castellani, V.; Bartimoccia, S.; Nocella, C.; Violi, F. Oleuropein, a component of extra virgin olive oil, lowers postprandial glycaemia in healthy subjects. Br. J. Clin. Pharmacol. 2018, 84, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Jemai, H.; Bouaziz, M.; Fki, I.; El Feki, A.; Sayadi, S. Hypolipidimic and antioxidant activities of oleuropein and its hydrolysis derivative-rich extracts from Chemlali olive leaves. Chem. Biol. Interact. 2008, 176, 88–98. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Barbosa, S.; Silva, M.; Monteiro, D.; Pinheiro, V.; Mourão, J.L.; Fernandes, J.; Rocha, S.; Belo, L.; Santos-Silva, A. The effect of olive leaf supplementation on the constituents of blood and oxidative stability of red blood cells. J. Funct. Food 2014, 9, 271–279. [Google Scholar] [CrossRef]
- Kim, S.W.; Hur, W.; Li, T.Z.; Lee, Y.K.; Choi, J.E.; Hong, S.W.; Lyoo, K.-S.; You, C.R.; Jung, E.S.; Jung, C.K.; et al. Oleuropein prevents the progression of steatohepatitis to hepatic fibrosis induced by a high-fat diet in mice. Exp. Mol. Med. 2014, 46, e92. [Google Scholar] [CrossRef]
- Porcu, C.; Sideri, S.; Martini, M.; Cocomazzi, A.; Galli, A.; Tarantino, G.; Balsano, C. Oleuropein Induces AMPK-Dependent Autophagy in NAFLD Mice, Regardless of the Gender. Int. J. Mol. Sci. 2018, 19, 3948. [Google Scholar] [CrossRef] [PubMed]
- Drira, R.; Chen, S.; Sakamoto, K. Oleuropein and hydroxytyrosol inhibit adipocyte differentiation in 3 T3-L1 cells. Life Sci. 2011, 89, 708–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svobodova, M.; Andreadou, I.; Skaltsounis, A.-L.; Kopecky, J.; Flachs, P. Oleuropein as an inhibitor of peroxisome proliferator-activated receptor gamma. Genes Nutr. 2014, 9, 376. [Google Scholar] [CrossRef] [PubMed]
- Malliou, F.; Andreadou, I.; Gonzalez, F.J.; Lazou, A.; Xepapadaki, E.; Vallianou, I.; Lambrinidis, G.; Mikros, E.; Marselos, M.; Skaltsounis, A.-L.; et al. The olive constituent oleuropein, as a PPARα agonist, markedly reduces serum triglycerides. J. Nutr. Biochem. 2018, 59, 17–28. [Google Scholar] [CrossRef]
- Kuem, N.; Song, S.J.; Yu, R.; Yun, J.W.; Park, T. Oleuropein attenuates visceral adiposity in high-fat diet-induced obese mice through the modulation of WNT10b- and galanin-mediated signalings. Mol. Nutr. Food Res. 2014, 58, 2166–2176. [Google Scholar] [CrossRef]
- Van der Stelt, I.; Hoek-van den Hil, E.F.; Swarts, H.J.M.; Vervoort, J.J.M.; Hoving, L.; Skaltsounis, L.; Lemonakis, N.; Andreadou, I.; van Schothorst, E.M.; Keijer, J. Nutraceutical oleuropein supplementation prevents high fat diet-induced adiposity in mice. J. Funct. Food 2015, 14, 702–715. [Google Scholar] [CrossRef]
- Oi-Kano, Y.; Iwasaki, Y.; Nakamura, T.; Watanabe, T.; Goto, T.; Kawada, T.; Watanabe, K.; Iwai, K. Oleuropein aglycone enhances UCP1 expression in brown adipose tissue in high-fat-diet-induced obese rats by activating β-adrenergic signaling. J. Nutr. Biochem. 2017, 40, 209–218. [Google Scholar] [CrossRef]
- Poudyal, H.; Campbell, F.; Brown, L. Olive Leaf Extract Attenuates Cardiac, Hepatic, and Metabolic Changes in High Carbohydrate–, High Fat–Fed Rats. J. Nutr. 2010, 140, 946–953. [Google Scholar] [CrossRef]
- Casamenti, F.; Grossi, C.; Rigacci, S.; Pantano, D.; Luccarini, I.; Stefani, M. Oleuropein Aglycone: A Possible Drug against Degenerative Conditions. In Vivo Evidence of its Effectiveness against Alzheimer’s Disease. J. Alzheimers Dis. 2015, 45, 679–688. [Google Scholar] [CrossRef]
- Yu, H.; Liu, P.; Tang, H.; Jing, J.; Lv, X.; Chen, L.; Jiang, L.; Xu, J.; Li, J. Oleuropein, a natural extract from plants, offers neuroprotection in focal cerebral ischemia/reperfusion injury in mice. Eur. J. Pharmacol. 2016, 775, 113–119. [Google Scholar] [CrossRef]
- Sun, W.; Wang, X.; Hou, C.; Yang, L.; Li, H.; Guo, J.; Huo, C.; Wang, M.; Miao, Y.; Liu, J.; et al. Oleuropein improves mitochondrial function to attenuate oxidative stress by activating the Nrf2 pathway in the hypothalamic paraventricular nucleus of spontaneously hypertensive rats. Neuropharmacology 2017, 113, 556–566. [Google Scholar] [CrossRef]
- Alirezaei, M.; Rezaei, M.; Hajighahramani, S.; Sookhtehzari, A.; Kiani, K. Oleuropein attenuates cognitive dysfunction and oxidative stress induced by some anesthetic drugs in the hippocampal area of rats. J. Physiol. Sci. 2017, 67, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Simsek, T.; Erbas, M.; Buyuk, B.; Pala, C.; Sahin, H.; Altinisik, B. Prevention of rocuronium induced mast cell activation with prophylactic oleuropein rich diet in anesthetized rabbits. Acta Cir. Bras. 2018, 33, 954–963. [Google Scholar] [CrossRef]
- Achour, I.; Arel-Dubeau, A.-M.; Renaud, J.; Legrand, M.; Attard, E.; Germain, M.; Martinoli, M.-G. Oleuropein Prevents Neuronal Death, Mitigates Mitochondrial Superoxide Production and Modulates Autophagy in a Dopaminergic Cellular Model. Int. J. Mol. Sci. 2016, 17, 1293. [Google Scholar] [CrossRef] [PubMed]
- Palazzi, L.; Bruzzone, E.; Bisello, G.; Leri, M.; Stefani, M.; Bucciantini, M.; Polverino de Laureto, P. Oleuropein aglycone stabilizes the monomeric α-synuclein and favours the growth of non-toxic aggregates. Sci. Rep. 2018, 8, 8337–8354. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Oropesa-Nuñez, R.; Canale, C.; Raimondi, S.; Giorgetti, S.; Bruzzone, E.; Bellotti, V.; Stefani, M.; Bucciantini, M. Oleuropein aglycone: A polyphenol with different targets against amyloid toxicity. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1432–1442. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wu, G.; Zou, X.; Jiang, K. Oleuropein protects intracerebral hemorrhage-induced disruption of blood-brain barrier through alleviation of oxidative stress. Pharmacol. Rep. 2017, 69, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, X.; Li, Q. Protective Effects of Oleuropein Against Cerebral Ischemia/Reperfusion by Inhibiting Neuronal Apoptosis. Med. Sci. Monit. 2018, 24, 6587–6598. [Google Scholar] [CrossRef]
- Lee, B.; Shim, I.; Lee, H.; Hahm, D.-H. Effect of oleuropein on cognitive deficits and changes in hippocampal brain-derived neurotrophic factor and cytokine expression in a rat model of post-traumatic stress disorder. J. Nat. Med. 2018, 72, 44–56. [Google Scholar] [CrossRef]
- Lee, B.; Shim, I.; Lee, H.; Hahm, D.-H. Oleuropein reduces anxiety-like responses by activating of serotonergic and neuropeptide Y (NPY)-ergic systems in a rat model of post-traumatic stress disorder. Anim. Cells Syst. 2018, 22, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Rabiei, Z.; Jahanbazi, S.; Alibabaei, Z. Antidepressant Effects of Oleuropein in Male Mice by Forced Swim Test and Tail Suspension Test. World Fam. Med. J. Middle East. J. Fam. Med. 2018, 16, 132–144. [Google Scholar] [CrossRef]
- Lee, O.-H.; Lee, B.-Y. Antioxidant and antimicrobial activities of individual and combined phenolics in Olea europaea leaf extract. Bioresour. Bioprocess. 2010, 101, 3751–3754. [Google Scholar] [CrossRef]
- Obied, H.K.; Bedgood, D.R.; Prenzler, P.D.; Robards, K. Bioscreening of Australian olive mill waste extracts: Biophenol content, antioxidant, antimicrobial and molluscicidal activities. Food Chem. Toxicol. 2007, 45, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Sudjana, A.N.; D’Orazio, C.; Ryan, V.; Rasool, N.; Ng, J.; Islam, N.; Riley, T.V.; Hammer, K.A. Antimicrobial activity of commercial Olea europaea (olive) leaf extract. J. Antimicrob. Agents 2009, 33, 461–463. [Google Scholar] [CrossRef]
- Lee-Huang, S.; Huang, P.L.; Zhang, D.; Lee, J.W.; Bao, J.; Sun, Y.; Chang, Y.-T.; Zhang, J.; Huang, P.L. Discovery of small-molecule HIV-1 fusion and integrase inhibitors oleuropein and hydroxytyrosol: Part, I. Integrase inhibition. Biochem. Biophys. Res. Commun. 2007, 354, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-Y.; Ima-Nirwana, S. Olives and Bone: A Green Osteoporosis Prevention Option. Int. J. Environ. Res. Public Health 2016, 13, 755. [Google Scholar] [CrossRef] [PubMed]
- Puel, C.; Mathey, J.; Agalias, A.; Kati-coulibaly, S.; Mardon, J.; Obled, C.; Davicco, M.-J.; Lebecque, P.; Horcajada, M.-N.; Skaltsounis, A.L.; et al. Dose–response study of effect of oleuropein, an olive oil polyphenol, in an ovariectomy/inflammation experimental model of bone loss in the rat. Clin. Nutr. 2006, 25, 859–868. [Google Scholar] [CrossRef]
- Santiago-Mora, R.; Casado-Díaz, A.; De Castro, M.D.; Quesada-Gómez, J.M. Oleuropein enhances osteoblastogenesis and inhibits adipogenesis: The effect on differentiation in stem cells derived from bone marrow. Osteoporos. Int. 2011, 22, 675–684. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Anter, J.; Müller, S.; Winter, P.; Quesada-Gómez, J.M.; Dorado, G. Transcriptomic analyses of the anti-adipogenic effects of oleuropein in human mesenchymal stem cells. Food Funct. 2017, 8, 1254–1270. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Li, X.; Lin, J.; Zheng, W.; Hu, Z.; Xuan, J.; Ni, W.; Pan, X. Oleuropein inhibits the IL-1β-induced expression of inflammatory mediators by suppressing the activation of NF-κB and MAPKs in human osteoarthritis chondrocytes. Food Funct. 2017, 8, 3737–3744. [Google Scholar] [CrossRef] [PubMed]
- Mehraein, F.; Sarbishegi, M.; Aslani, A. Therapeutic Effects of Oleuropein on Wounded Skin in Young Male Balb/c Mice. Wounds 2014, 26, 7. [Google Scholar]
- Bharathy, H.; Fathima, N.N. Exploiting oleuropein for inhibiting collagen fibril formation. Int. J. Biol. Macromol. 2017, 101, 179–186. [Google Scholar] [CrossRef]
- Ji, S.T.; Kim, Y.-J.; Jung, S.Y.; Kim, D.Y.; Kang, S.; Park, J.H.; Jang, W.B.; Ha, J.; Yun, J.; Kwon, S.-M. Oleuropein attenuates hydrogen peroxide-induced autophagic cell death in human adipose-derived stem cells. Biochem. Biophys. Res. Commun. 2018, 499, 675–680. [Google Scholar] [CrossRef]
- Margheri, F.; Menicacci, B.; Laurenzana, A.; Del Rosso, M.; Fibbi, G.; Cipolleschi, M.G.; Ruzzolini, J.; Nediani, C.; Mocali, A.; Giovannelli, L. Oleuropein aglycone attenuates the pro-angiogenic phenotype of senescent fibroblasts: A functional study in endothelial cells. J. Funct. Food 2019, 53, 219–226. [Google Scholar] [CrossRef]
- Vissers, M.N.; Zock, P.L.; Roodenburg, A.J.C.; Leenen, R.; Katan, M.B. Olive Oil Phenols Are Absorbed in Humans. J. Nutr. 2002, 132, 409–417. [Google Scholar] [CrossRef]
- Gikas, E.; Papadopoulos, N.; Tsarbopoulos, A. Kinetic Study of the Acidic Hydrolysis of Oleuropein, the Major Bioactive Metabolite of Olive Oil. J. Liq. Chromatogr. Relat. Technol. 2006, 29, 497–508. [Google Scholar] [CrossRef]
- Carrera-González, M.P.; Ramírez-Expósito, M.J.; Mayas, M.D.; Martínez-Martos, J.M. Protective role of oleuropein and its metabolite hydroxytyrosol on cancer. Trends Food Sci. Technol. 2013, 31, 92–99. [Google Scholar] [CrossRef]
- Briante, R.; Patumi, M.; Terenziani, S.; Bismuto, E.; Febbraio, F.; Nucci, R. Olea europaea L. Leaf Extract and Derivatives: Antioxidant Properties. J. Agric. Food Chem. 2002, 50, 4934–4940. [Google Scholar] [CrossRef]
- Beauchamp, G.K.; Keast, R.S.J.; Morel, D.; Lin, J.; Pika, J.; Han, Q.; Lee, C.-H.; Smith, A.B.; Breslin, P.A.S. Ibuprofen-like activity in extra-virgin olive oil: Phytochemistry. Nature 2005, 437, 45–46. [Google Scholar] [CrossRef]
- Iacono, A.; Gómez, R.; Sperry, J.; Conde, J.; Bianco, G.; Meli, R.; Gómez-Reino, J.J.; Smith, A.B.; Gualillo, O. Effect of oleocanthal and its derivatives on inflammatory response induced by lipopolysaccharide in a murine chondrocyte cell line. Arthritis Rheum. 2010, 62, 1675–1682. [Google Scholar] [CrossRef]
- Scotece, M.; Conde, J.; Abella, V.; Lopez, V.; Francisco, V.; Ruiz, C.; Campos, V.; Lago, F.; Gomez, R.; Pino, J.; et al. Oleocanthal Inhibits Catabolic and Inflammatory Mediators in LPS-Activated Human Primary Osteoarthritis (OA) Chondrocytes Through MAPKs/NF-kappa B Pathways. Cell Phys. Biochem. 2018, 49, 2414–2426. [Google Scholar] [CrossRef]
- Segura Palacios, J.M.; Blazquez Sanchez, N.; Rivas Ruiz, F.; Aguilar Bernier, M.; Ramirez Lopez, B.; Fernandez Sanchez, M.E.; de Troya Martin, M. Topical treatment with oleocanthal extract in reducing inflammatory reactions after photodynamic therapy: A prospective quasi-experimental pilot study. Complement. Med. Res. 2019, 42, 298–301. [Google Scholar] [CrossRef]
- Agrawal, K.; Melliou, E.; Li, X.; Pedersen, T.L.; Wang, S.C.; Magiatis, P.; Newman, J.W.; Holt, R.R. Oleocanthal-rich extra virgin olive oil demonstrates acute anti-platelet effects in healthy men in a randomized trial. J. Funct. Food 2017, 36, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Abuznait, A.H.; Qosa, H.; Busnena, B.A.; El Sayed, K.A.; Kaddoumi, A. Olive-Oil-Derived Oleocanthal Enhances β-Amyloid Clearance as a Potential Neuroprotective Mechanism against Alzheimer’s Disease: In Vitro and in Vivo Studies. ACS Chem. Neurosci. 2013, 4, 973–982. [Google Scholar] [CrossRef]
- Parkinson, L.; Keast, R. Oleocanthal, a Phenolic Derived from Virgin Olive Oil: A Review of the Beneficial Effects on Inflammatory Disease. Int. J. Mol. Sci. 2014, 15, 12323–12334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batarseh, Y.S.; Mohamed, L.A.; Al Rihani, S.B.; Mousa, Y.M.; Siddique, A.B.; El Saved, K.A.; Kaddoumi, A. Oleocanthal ameliorates amyloid-beta oligomers’ toxicity on astrocytes and neuronal cells: In vitro studies. Neuroscience 2017, 352, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Giusti, L.; Angeloni, C.; Barbalace, M.C.; Lacerenza, S.; Ciregia, F.; Ronci, M.; Urbani, A.; Manera, C.; Digiacomo, M.; Macchia, M.; et al. A Proteomic Approach to Uncover Neuroprotective Mechanisms of Oleocanthal against Oxidative Stress. Int. J. Mol. Sci. 2018, 19, 2329. [Google Scholar] [CrossRef] [PubMed]
- Qosa, H.; Batarseh, Y.S.; Mohyeldin, M.M.; El Sayed, K.A.; Keller, J.N.; Kaddoumi, A. Oleocanthal Enhances Amyloid-beta Clearance from the Brains of TgSwDI Mice and in Vitro across a Human Blood-Brain Barrier Model. ACS Chem. Neurosci. 2015, 6, 1849–1859. [Google Scholar] [CrossRef]
- Khanal, P.; Oh, W.-K.; Yun, H.J.; Namgoong, G.M.; Ahn, S.-G.; Kwon, S.-M.; Choi, H.-K.; Choi, H.S. p-HPEA-EDA, a phenolic compound of virgin olive oil, activates AMP-activated protein kinase to inhibit carcinogenesis. Carcinogenesis 2011, 32, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Elnagar, A.; Sylvester, P.; El Sayed, K. (−)-Oleocanthal as a c-Met Inhibitor for the Control of Metastatic Breast and Prostate Cancers. Planta Med. 2011, 77, 1013–1019. [Google Scholar] [CrossRef]
- Akl, M.R.; Ayoub, N.M.; Mohyeldin, M.M.; Busnena, B.A.; Foudah, A.I.; Liu, Y.-Y.; Sayed, K.A.E. Olive Phenolics as c-Met Inhibitors: (−)-Oleocanthal Attenuates Cell Proliferation, Invasiveness, and Tumor Growth in Breast Cancer Models. PLoS ONE 2014, 9, e97622. [Google Scholar] [CrossRef] [PubMed]
- Busnena, B.A.; Foudah, A.I.; Melancon, T.; El Sayed, K.A. Olive secoiridoids and semisynthetic bioisostere analogues for the control of metastatic breast cancer. Bioorg. Med. Chem. 2013, 21, 2117–2127. [Google Scholar] [CrossRef] [PubMed]
- Mohyeldin, M.M.; Akl, M.R.; Ebrahim, H.Y.; Dragoi, A.M.; Dykes, S.; Cardelli, J.A.; Sayed, K.A.E. The oleocanthal-based homovanillyl sinapate as a novel c-Met inhibitor. Oncotarget 2016, 7, 32247–32273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanfar, M.A.; Bardaweel, S.K.; Akl, M.R.; El Sayed, K.A. Olive Oil-derived Oleocanthal as Potent Inhibitor of Mammalian Target of Rapamycin: Biological Evaluation and Molecular Modeling Studies: Oleocanthal Is a Potent mTOR Inhibitor. Phytother. Res. 2015, 29, 1776–1782. [Google Scholar] [CrossRef]
- Ayoub, N.M.; Siddique, A.B.; Ebrahim, H.Y.; Mohyeldin, M.M.; El Sayed, K.A. The olive oil phenolic (-)-oleocanthal modulates estrogen receptor expression in luminal breast cancer in vitro and in vivo and synergizes with tamoxifen treatment. Eur. J. Pharmacol. 2017, 810, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Diez-Bello, R.; Jardin, I.; Lopez, J.J.; El Haouari, M.; Ortega-Vidal, J.; Altarejos, J.; Salido, G.M.; Salido, S.; Rosado, J.A. (-)-Oleocanthal inhibits proliferation and migration by modulating Ca2+ entry through TRPC6 in breast cancer cells. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Fogli, S.; Arena, C.; Carpi, S.; Polini, B.; Bertini, S.; Digiacomo, M.; Gado, F.; Saba, A.; Saccomanni, G.; Breschi, M.C.; et al. Cytotoxic Activity of Oleocanthal Isolated from Virgin Olive Oil on Human Melanoma Cells. Nutr. Cancer 2016, 68, 873–877. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, J.; Peng, L. (−)-Oleocanthal exerts anti-melanoma activities and inhibits STAT3 signaling pathway. Oncol. Rep. 2017, 37, 483–491. [Google Scholar] [CrossRef]
- Polini, B.; Digiacomo, M.; Carpi, S.; Bertini, S.; Gado, F.; Saccomanni, G.; Macchia, M.; Nieri, P.; Manera, C.; Fogli, S. Oleocanthal and oleacein contribute to the in vitro therapeutic potential of extra virgin oil-derived extracts in non-melanoma skin cancer. Oxicol. In Vitro 2018, 52, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Pei, T.; Meng, Q.; Han, J.; Sun, H.; Li, L.; Song, R.; Sun, B.; Pan, S.; Liang, D.; Liu, L. (-)-Oleocanthal inhibits growth and metastasis by blocking activation of STAT3 in human hepatocellular carcinoma. Oncotarget. 2016, 7, 43475–43491. [Google Scholar] [CrossRef]
- Cusimano, A.; Balasus, D.; Azzolina, A.; Augello, G.; Emma, M.R.; Di Sano, C.; Gramignoli, R.; Strom, S.C.; McCubery, J.A.; Montalto, G.; et al. Oleocanthal exerts antitumor effects on human liver and colon cancer cells through ROS generation. Int. J. Oncol. 2017, 51, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Scotece, M.; Gómez, R.; Conde, J.; Lopez, V.; Gómez-Reino, J.J.; Lago, F.; Smith, A.B.; Gualillo, O. Oleocanthal Inhibits Proliferation and MIP-1 Expression in Human Multiple Myeloma Cells. Curr. Med. Chem. 2013, 20, 2467–2475. [Google Scholar] [CrossRef]
- Pang, K.-L.; Chin, K.-Y. The Biological Activities of Oleocanthal from a Molecular Perspective. Nutrients 2018, 10, 570. [Google Scholar] [CrossRef]
- García-Villalba, R.; Carrasco-Pancorbo, A.; Oliveras-Ferraros, C.; Vázquez-Martín, A.; Menéndez, J.A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Characterization and quantification of phenolic compounds of extra-virgin olive oils with anticancer properties by a rapid and resolutive LC-ESI-TOF MS method. J. Pharm. Biomed. Anal. 2010, 51, 416–429. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Santos, V.; Mangericão, H.; Gordon, M.H. Effects of Copper on the Antioxidant Activity of Olive Polyphenols in Bulk Oil and Oil-in-Water Emulsions. J. Agric. Food Chem. 2006, 54, 3738–3743. [Google Scholar] [CrossRef]
- Czerwińska, M.; Kiss, A.K.; Naruszewicz, M. A comparison of antioxidant activities of oleuropein and its dialdehydic derivative from olive oil, oleacein. Food Chem. 2012, 131, 940–947. [Google Scholar] [CrossRef]
- Naruszewicz, M.; Czerwinska, M.E.; Kiss, A.K. Oleacein. Translation from Mediterranean Diet to Potential Antiatherosclerotic Drug. Curr. Pharm. Des. 2015, 21, 1205–1212. [Google Scholar] [CrossRef]
- Filipek, A.; Czerwinska, M.E.; Kiss, A.K.; Wrzosek, M.; Naruszewicz, M. Oleacein enhances anti-inflammatory activity of human macrophages by increasing CD163 receptor expression. Phytomedicine 2015, 22, 1255–1261. [Google Scholar] [CrossRef]
- Segade, M.; Bermejo, R.; Silva, A.; Paiva-Martins, F.; Gil-Longo, J.; Campos-Toimil, M. Involvement of endothelium in the vasorelaxant effects of 3,4-DHPEA-EA and 3,4-DHPEA-EDA, two major functional bioactives in olive oil. J. Funct. Food 2016, 23, 637–646. [Google Scholar] [CrossRef]
- Filipek, A.; Czerwinska, M.E.; Kiss, A.K.; Polanski, J.A.; Naruszewicz, M. Oleacein may inhibit destabilization of carotid plaques from hypertensive patients. Impact on high mobility group protein-1. Phytomedicine 2017, 32, 68–73. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Kiss, A.K.; Naruszewicz, M. Inhibition of human neutrophils NEP activity, CD11b/CD18 expression and elastase release by 3,4-dihydroxyphenylethanol-elenolic acid dialdehyde, oleacein. Food Chem. 2014, 153, 1–8. [Google Scholar] [CrossRef]
- Lombardo, G.E.; Lepore, S.M.; Morittu, V.M.; Arcidiacono, B.; Colica, C.; Procopio, A.; Maggisano, V.; Bulotta, S.; Costa, N.; Mignogna, C.; et al. Effects of Oleacein on High-Fat Diet-Dependent Steatosis, Weight Gain, and Insulin Resistance in Mice. Front. Endocrinol. 2018, 9, 116. [Google Scholar] [CrossRef] [Green Version]
- Cicero, A.F.G.; Nascetti, S.; López-Sabater, M.C.; Elosua, R.; Salonen, J.T.; Nyyssönen, K.; Poulsen, H.E.; Zunft, H.J.; Kiesewetter, H.; de la Torre, K.; et al. Changes in LDL fatty acid composition as a response to olive oil treatment are inversely related to lipid oxidative damage: The EUROLIVE study. J. Am. Coll. Nutr. 2008, 27, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage (ID 1333, 1638, 1639, 1696, 2865), maintenance of normal blood HDL cholesterol concentrations (ID 1639), mainte. EFSA J. 2011, 9, 2033. [CrossRef]
- Valls, R.-M.; Farràs, M.; Suárez, M.; Fernández-Castillejo, S.; Fitó, M.; Konstantinidou, V.; Fuentes, F.; López-Miranda, J.; Giralt, M.; Covas, M.; et al. Effects of functional olive oil enriched with its own phenolic compounds on endothelial function in hypertensive patients. A randomised controlled trial. Food Chem. 2015, 167, 30–35. [Google Scholar]
- Hohmann, C.D.; Cramer, H.; Michalsen, A.; Kessler, C.; Steckhan, N.; Choi, K.; Dobos, G. Effects of high phenolic olive oil on cardiovascular risk factors: A systematic review and meta-analysis. Phytomedicine 2015, 22, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Buckland, G.; Travier, N.; Agudo, A.; Fonseca-Nunes, A.; Navarro, C.; Lagiou, P.; Demetriou, C.; Amiano, P.; Dorronsoro, M.; Chirlaque, M.D.; et al. Olive oil intake and breast cancer risk in the Mediterranean countries of the European Prospective Investigation into Cancer and Nutrition study. Int. J. Cancer 2012, 131, 2465–2469. [Google Scholar] [CrossRef] [Green Version]
- Medina-Remon, A.; Casas, R.; Tressserra-Rimbau, A.; Ros, E.; Martinez-Gonzalez, M.A.; Fito, M.; Corella, D.; Salas-Salvadó, J.; Lamuela-Raventos, R.M.; Estruch, R. Polyphenol intake from a Mediterranean diet decreases inflammatory biomarkers related to atherosclerosis: A substudy of the PREDIMED trial. Bri. J. Clin. Pharmacol. 2017, 83, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Toledo, E.; Salas-Salvadó, J.; Donat-Vargas, C.; Buil-Cosiales, P.; Estruch, R.; Ros, E.; Corella, D.; Fitó, M.; Hu, F.B.; Arós, F.; et al. Mediterranean diet and invasive breast cancer risk among women at high cardiovascular risk in the PREDIMED trial. JAMA Intern. Med. 2015, 175, 1752–1760. [Google Scholar] [CrossRef]
- Valls-Pedret, C.; Sala-Vila, A.; Serra-Mir, M.; Corella, D.; de la Torre, R.; Martínez-González, M.Á.; Martínez-Lapiscina, E.H.; Fitó, M.; Pérez-Heras, A.; Salas-Salvadó, J.; et al. Mediterranean diet and age-related cognitive decline. JAMA Intern. Med. 2015, 175, 1094–1103. [Google Scholar] [CrossRef]
- Martínez-Lapiscina, E.H.; Clavero, P.; Toledo, E.; San Julián, B.; Sanchez-Tainta, A.; Corella, D.; Lamuela-Raventós, R.M.; Martínez, J.A.; Martínez-Gonzalez, M.Á. Virgin olive oil supplementation and long-term cognition: The PREDIMEDNAVARRA randomized, trial. J. Nutr. Health Aging. 2013, 17, 544–552. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02842567 (accessed on 13 May 2019).
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02068092 (accessed on 13 May 2019).






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karković Marković, A.; Torić, J.; Barbarić, M.; Jakobušić Brala, C. Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health. Molecules 2019, 24, 2001. https://doi.org/10.3390/molecules24102001
Karković Marković A, Torić J, Barbarić M, Jakobušić Brala C. Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health. Molecules. 2019; 24(10):2001. https://doi.org/10.3390/molecules24102001
Chicago/Turabian StyleKarković Marković, Ana, Jelena Torić, Monika Barbarić, and Cvijeta Jakobušić Brala. 2019. "Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health" Molecules 24, no. 10: 2001. https://doi.org/10.3390/molecules24102001
APA StyleKarković Marković, A., Torić, J., Barbarić, M., & Jakobušić Brala, C. (2019). Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health. Molecules, 24(10), 2001. https://doi.org/10.3390/molecules24102001

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}