- 1Microbiology & Molecular Genetics Department, Faculty of Medicine, The Hebrew University of Jerusalem, Jerusalem, Israel
1耶路撒冷希伯来大学医学院微生物学与分子遗传学系,以色列耶路撒冷 - 2The Rachel and Selim Benin School of Computer Science and Engineering, The Hebrew University of Jerusalem, Jerusalem, Israel
阿拉伯数字以色列耶路撒冷希伯来大学 Rachel and Selim Benin 计算机科学与工程学院
Human milk oligosaccharides (HMOs) are a family of glycans found in breastmilk with over 200 identified structures. Despite being the third-largest solid component in breastmilk, HMOs are indigestible by infants, and they serve as food for the infant gut bacteria. Most research thus far has focused on Bifidobacterium species that harbor many glycoside hydrolases (GHs) tailored to break the carbon bonds in HMO molecules. However, there are additional microbes in the infant gut, such as Bacteroides species, with increasing evidence that they, too, are able to break-down HMOs. To study the unbiased impact of breastfeeding on the infant gut microbiome, we need to investigate the underlying mechanisms of HMO utilization by all members of the infant gut. Here, we developed an optimized system for isolating Bacteroides strains from infant stool samples. We then examined the HMO utilization capacity of multiple Bacteroides isolates by performing growth curves on six common HMOs (2’-FL, DFL, 3’-SL, 6’-SL, LNT, LNnT). Isolates often displayed similar growth characteristics on similarly-structured HMOs, like sialylated or fucosylated sugars. We identified variation in HMO utilization across multiple strains of the same species, and chose to focus here on a Bacteroides dorei isolate that was able to utilize the test HMOs. We performed RNA sequencing on B. dorei cultures, comparing the transcriptional profile in minimal media supplemented with glucose or HMOs. We showed that B. dorei employs an extensive metabolic response to HMOs. Surprisingly, there was no clear up-regulation for most GH families previously known to break-down HMOs, possibly because they were almost exclusively described in Bifidobacterium species. Instead, B. dorei exhibits a generalized response to HMOs, markedly up-regulating several shared GH families across all conditions. Within each GH family, B. dorei displays a consistent pattern of up-regulation of some genes with down-regulation of the others. This response pattern to HMOs has yet to be described in other commensals of the infant gut. Our work highlights the importance of expanding the HMO-microbiome studies beyond Bifidobacterium species, sheds light on the differences across Bacteroides strains in terms of HMO utilization, and paves the way to understanding the mechanisms enabling Bacteroides HMO utilization.
母乳低聚糖 (HMO) 是母乳中发现的聚糖家族,具有 200 多种已鉴定的结构。尽管 HMO 是母乳中的第三大固体成分,但婴儿难以消化,它们是婴儿肠道细菌的食物。迄今为止,大多数研究都集中在双歧杆菌物种上,这些细菌含有许多糖苷水解酶 (GH),这些水解酶 (GH) 是为了打破 HMO 分子中的碳键而定制的。然而,婴儿肠道中还有其他微生物,例如 拟杆菌属,越来越多的证据表明它们也能够分解 HMO。为了研究母乳喂养对婴儿肠道微生物组的公正影响,我们需要调查婴儿肠道所有成员利用 HMO 的潜在机制。在这里,我们开发了一种优化的系统,用于从婴儿粪便样本中分离拟杆菌菌株。然后,我们通过在 6 种常见 HMO (2'-FL、DFL、3'-SL、6'-SL、LNT、LNnT) 上执行生长曲线来检查多种拟杆菌属分离株的 HMO 利用能力。分离株通常在结构相似的 HMO 上表现出相似的生长特性,如唾液酸化或岩藻糖基化糖。我们确定了同一物种的多个菌株中 HMO 利用率的差异,并选择在这里专注于能够利用测试 HMO 的 Bacteroides dorei 分离株。我们对 B. dorei 培养物进行了 RNA 测序,比较了添加葡萄糖或 HMO 的最小培养基中的转录谱。我们表明 B. dorei 对 HMO 具有广泛的代谢反应。 令人惊讶的是,对于以前已知的大多数分解 HMO 的 GH 家族,没有明显的上调,可能是因为它们几乎只在双歧杆菌物种中被描述。相反,B. dorei 表现出对 HMO 的普遍反应,在所有情况下显着上调几个共享的 GH 家族。在每个 GH 家族中,B. dorei 表现出一些基因上调而其他基因下调的一致模式。这种对 HMO 的反应模式尚未在婴儿肠道的其他共生体中描述。我们的工作强调了将 HMO 微生物组研究扩展到双歧杆菌物种之外的重要性,阐明了拟杆菌菌株在 HMO 利用方面的差异,并为理解使拟杆菌 HMO 利用的机制铺平了道路。
Introduction 介绍
The infant gut microbiome is very dynamic (Yassour et al., 2016; Ferretti et al., 2018), and stabilizes to an adult-like state around the age of three (Yatsunenko et al., 2012; Yassour et al., 2016). The initial colonization of the infant gut is a complex process (La Rosa et al., 2014; Lax et al., 2014), mainly influenced by delivery mode (Dominguez-Bello et al., 2010; Azad et al., 2013; Madan et al., 2016; Yassour et al., 2016) and infant feeding (formula vs. breast milk) (Azad et al., 2013; Bäckhed et al., 2015; Madan et al., 2016; Stewart et al., 2018). Human Milk Oligosaccharides (HMOs) are a family of glycans found in breast milk, and despite being the third-largest solid component in human milk (Vandenplas et al., 2018), infants themselves are unable to digest these sugars (Engfer et al., 2000; Gnoth et al., 2000), and HMOs are exclusively digested by bacteria in the infant gut. HMOs take part in many important processes, such as infant immune system maturation (Plaza-Díaz et al., 2018) and brain development (Jacobi et al., 2016; Hegar et al., 2019), but one of their most-studied roles is the promotion of a healthy infant gut microbial community, by supporting the colonization of beneficial gut bacteria that can break down the HMO chemical structure (Walsh et al., 2020). Mounting evidence show that early establishment of a “correct” microbial community in the infant gut is crucial for future health, in microbiota-mediated colonization resistance against intestinal pathogens (Buffie and Pamer, 2013; McLaren and Callahan, 2020), even under antibiotic disturbance (Letten et al., 2021a), and the ability of the microbiome to modulate persistence of antibiotic resistant strains (Letten et al., 2021b). The importance of HMOs in shaping the unique infant microbiome has been a subject of avid research in the past two decades, however most of it focused on Bifidobacterium species (Dethlefsen et al., 2007; Sela et al., 2008).
婴儿肠道微生物组非常动态(Yassour et al., 2016;Ferretti等 人,2018 年),并在三岁左右稳定到类似成人的状态(Yatsunenko等 人,2012 年;Yassour等 人,2016 年)。婴儿肠道的初始定植是一个复杂的过程(La Rosa等 人,2014 年;Lax et al., 2014),主要受交付模式的影响(Dominguez-Bello et al., 2010;Azad等 人,2013 年;Madan等 人,2016 年;Yassour等 人,2016 年)和婴儿喂养(配方奶粉与母乳)(Azad et al., 2013;Bäckhed等 人,2015 年;Madan等 人,2016 年;Stewart et al., 2018)。母乳低聚糖 (HMO) 是母乳中发现的聚糖家族,尽管它是母乳中的第三大固体成分(Vandenplas 等 人,2018 年),但婴儿本身无法消化这些糖(Engfer 等 人,2000 年;Gnoth 等 人,2000 年),而 HMO 仅被婴儿肠道中的细菌消化。HMO 参与许多重要过程,例如婴儿免疫系统成熟(Plaza-Díaz et al., 2018)和大脑发育(Jacobi et al., 2016;Hegar et al., 2019),但他们研究最多的作用之一是通过支持可以分解 HMO 化学结构的有益肠道细菌的定植来促进健康的婴儿肠道微生物群落(Walsh et al., 2020)。 越来越多的证据表明,在微生物群介导的对肠道病原体的定植抵抗力中,在婴儿肠道中及早建立“正确”的微生物群落对未来的健康至关重要(Buffie 和 Pamer,2013 年;McLaren 和 Callahan,2020 年),即使在抗生素干扰下(Letten等 人,2021a),以及微生物组调节抗生素耐药菌株持久性的能力(Letten 等人 ,2021b)。在过去的二十年里,HMO 在塑造独特的婴儿微生物组方面的重要性一直是狂热研究的主题,但其中大部分集中在双歧杆菌物种上(Dethlefsen 等 人,2007 年;Sela et al., 2008)。
Bifidobacterium species abundance is strongly associated with HMO consumption, as demonstrated by multiple studies comparing breastfed to non-breastfed infants (Davis et al., 2016; Smith-Brown et al., 2016; Bai et al., 2018; Lawson et al., 2020; Laursen et al., 2021; Liu et al., 2021). The first HMO-specific utilization cluster (H1) was discovered in 2008 (Sela et al., 2008). It is 43kb long and is conserved in all B. longum subsp. infantis genomes sequenced thus far. Since 2008, five additional HMO-utilizing gene clusters (H2, H3, H4, H5, urease) have been identified (LoCascio et al., 2010), albeit they are less conserved. Recently, additional genes capable of HMO utilization were discovered, which do not necessarily belong to the known clusters identified thus far, and revealed variation in differential expression of genes across Bifidobacterium longum subsp. infantis bacteria (Zabel et al., 2020). Importantly, none of these well-studied gene clusters were found in any non-Bifidobacterium species (Sela and Mills, 2010; Marcobal and Sonnenburg, 2012).
Despite the well-established connection between Bifidobacterium and HMOs, recent empirical evidence support the HMO utilization by other, non-Bifidobacterium species. First, some breastfeeding infants do not harbor any Bifidobacterium species throughout their breastfeeding period (Yassour et al., 2016), and findings exhibit inter-study heterogeneity (Harmsen et al., 2000; Favier et al., 2002; Palmer et al., 2007). Importantly, it was recently reported that the best known HMO-utilizer strain, Bifidobacterium longum subsp. infantis, considered to be predominant in the infant gut (Chichlowski et al., 2020), is not evident in 90% of infants in the US (Casaburi et al., 2021) and Finland (Vatanen et al., 2019). Second, there are significant correlations between specific HMO levels and the relative abundance of other genera, such as Lactobacillus (Bai et al., 2018), Bacteroides and Parabacteroides (Wang et al., 2015; Borewicz et al., 2019; Borewicz et al., 2020). Specifically, the immediate next candidates for HMO utilization are Bacteroides species, which are known degraders of other complex versatile glycans (Hooper et al., 2002; Bjursell et al., 2006; Martens et al., 2008), like mucin coating the colon epithelium (Marcobal et al., 2011; Pruss et al., 2021). When examining the genomes of common infant gut microbes, we can computationally annotate some genes as potential HMO-utilizers (Xu et al., 2003; Yassour et al., 2016; Vatanen et al., 2019), mostly from the Bacteroides genus. Lastly, there is experimental evidence that various Bacteroides strains can utilize HMOs as their sole carbon source (Salli et al., 2021), and some of the mechanisms have been further characterized (Marcobal et al., 2011). Bacteroides thetaiotaomicron and Bacteroides fragilis utilized both host mucus and HMOs, using a similar set of upregulated genes (Marcobal et al., 2011). The variety of findings concerning non-Bifidobacterium HMO-degraders highlight the importance of expanding our knowledge of HMO utilization among other gut commensals.
We therefore place Bacteroides at the heart of this research. First, we developed an optimized system for isolation of Bacteroides strains from infant stool samples, utilizing Bacteroides-specific qPCR and Bacteroides-selective media. Second, we examined the HMO utilization capacity of our Bacteroides isolates by analyzing growth curves on six common HMO molecules: 2-Fucosyllactose (2’-FL), Difucosyllactose (DFL), 3-Sialyllactose (3’-SL), 6-Sialyllactose (6’-SL), Lacto-N-Tetraose (LNT) and Lacto-N-neotetraose (LNnT). Isolates often displayed similar growth characteristics on similarly-structured HMOs, like sialylated or fucosylated sugars. We identified variation in HMO utilization across multiple strains of the same species, and chose to focus here on a Bacteroides dorei isolate that was able to utilize the test HMOs. We performed RNA sequencing on B. dorei cultures, comparing the transcriptional profile in minimal media supplemented with glucose or HMO. We showed that B. dorei employs an extensive metabolic response to HMOs: 17 GH families were upregulated when grown on 2’-FL, 21 on DFL, 19 on 3’-SL, 23 on 6’-SL, 15 on LNT, and 18 on LNnT. In addition, B. dorei exhibits a generalized response to HMOs, markedly up-regulating several shared GH families across all conditions, in contrast to Bifidobacterium species, previously shown to employ specific GH families under different growth conditions. Within each GH family, B. dorei displays a consistent pattern of up-regulation of some genes with down-regulation of the others.
Materials and Methods
Bacteroides Isolation From Stool Samples
Ten stool samples were collected from seven exclusively breastfed infants less than six month old (S4-5, S5-6, S9-10 were collected twice from the same infants with an average of one week interval). Infants were vaginally-born, and did not receive any antibiotic treatment from birth to sample collection time. All seven mothers have agreed to participate in our study, which was approved by the Hebrew University’s Institutional Review Board (IRB), and signed our consent forms. Samples were collected from diapers using a small plastic spoon and tube, stored at 4°C, then transferred to -80°C within 8 hours. The samples were frozen for a maximal time of two months, and thawed twice: once for DNA extraction, and once for plating and Bacteroides isolation.
DNA was extracted using the DNeasy PowerSoil Pro Kit (QIAGEN) and used as a template for quantitative PCR (qPCR) reactions, with both general 16S primers (Ivanov et al., 2009; Yassour et al., 2018) (16S_F 5′ GGTGAATACGTTCCCGG 3′, 16S_R 5′ TACGGCTACCTTGTTACGACTT 3′) and Bacteroides-specific primers (Matsuki et al., 2002) (g-Bfra-F 5′ ATAGCCTTTCGAAAGRAAGAT 3′, g-Bfra-R 5′ CCAGTATCAACTGCAATTTTA 3′). qPCR reaction volumes were 1 μL 2.5 ng/μL DNA, 0.25 μL 10μM forward primer, 0.25 μL 10μM reverse primer, 3.5 μL Nuclease-free water, 5 μL SYBR Green PCR Master Mix. The amplification program for the Bacteroides-specific qPCR consisted of (1) 94°C for 5 min, (2) 94°C for 20s (3) 50°C for 20 s (4) 72°C for 50 s (5) repeat 2 - 4 for 39 times (6) 94°C for 15 s. The amplification program for the general 16S qPCR consisted of (1) 95°C for 10 min, (2) 95°C for 15 s, (3) 60°C for 30 s, and (4) repeat 2 and 3 39 times. The fluorescent products were detected at the last step of each cycle, and reactions were conducted on a BioRad CFX96 Real-Time System.
The cycle threshold (Ct) value was recorded for each sample, and it is inversely proportional to the amount of Bacteroides DNA in the sample. The negative control samples (NC1, NC2), that did not contain Bacteroides according to previously obtained in-lab 16S metagenomic sequencing, had a high Ct in Bacteroides-specific qPCR experiments. In contrast, DNA extracted from a pure Bacteroides fragilis (ATCC 25285) culture, serving as a positive control, had a low Ct value.
Samples that passed the initial qPCR screening were plated on blood agar plates (hylabs, PD005) and incubated at 37°C for 48 hours in an anaerobic chamber (COY). Then, colonies were re-streaked onto Bile Esculin Agar (BEA) plates and incubated in the same conditions. PCR reaction for the 16S region was performed using the universal primers 27F AGAGTTTGATCMTGGCTCAG, 1492R GGTTACCTTGTTACGACTT (Lane, 1991). Reaction volumes were 1 μL DNA, 1.25 μL 10μM forward primer, 1.25 μL 10μM reverse primer, 9 μL Nuclease-free water, 12.5 μL SYBR NEB Q5 High-Fidelity MasterMix. The program consisted of (1) 98°C for 30 s, (2) 98°C for 10s (3) 55°C for 15 s (4) 72°C for 30 s (5) repeat 2 - 4 for 29 times (6) 72°C for 2 min. AMPure XP (Beckman Coulter, A63881) bead cleaning of PCR products was performed according to manufacturer’s instructions. Then, Sanger sequencing of the 16S region was performed with the primer 515F GTGCCAGCMGCCGCGGTAA (Caporaso et al., 2011), at Hylabs (Rehovot, Israel). Bacterial identity identification was achieved using BLAST (Altschul et al., 1990) alignment with default settings.
16S Ribosomal Gene Sequencing of Stool Samples
Samples that were found to contain Bacteroides according to qPCR screening were chosen for 16S rRNA sequencing. 16S rRNA gene sequencing was performed as previously described (Caporaso et al., 2012), and the 16S gene data set consists of sequences targeting the V4 variable region. Sequencing was performed on the Illumina MiSeq platform according to the manufacturer’s specifications, with addition of 5% PhiX, generating single-end reads of 250 bp in length. Human reads were excluded from further processing with Bowtie2 (Langmead and Salzberg, 2012), in addition to quality-based filtering of reads performed using Fastqmcf (Aronesty, 2013). Taxonomic classification was assigned using BURST (Al-Ghalith and Knights, 2020).
Growth Curves
For each growth curve experiment, an isolate from infant stool samples was plated on a brain heart infusion (BHI) agar plate (Sigma 70138) supplemented with 50 ml/L fetal bovine serum (FBS; Sigma F2442), 10 ml/L trace vitamins (ATCC® MD-VS™), 10 ml/L trace minerals (ATCC® MD-TMS™), 10 ml/L vitamin K1 and hemin (BBL, 212354), 1 g/L D-(+)-Cellobiose (Alfa Aesar, 528507), 1 g/L D-(+)-Maltose (Caisson, 6363537), 1 g/L D-(+)-Fructose (Sigma Aldrich, 1286504), and 0.5 g/L L-Cysteine (Acros Organics, 52904). The individual HMOs were received as a donation from the DSM Nutritional Products Ltd (previously known as Glycom).
The plates were incubated in an anaerobic chamber (COY) for 48 hours, at 37°C. Two single colonies (biological replicates) were transferred to liquid BHI media (Sigma 53286), supplemented as described above, for overnight incubation. These cultures were then diluted 1:100 in Bacteroides-specific minimal-media (MM) with 0.5% (w/v) glucose (Martens et al., 2008). After another overnight incubation, the culture was diluted 1:100 into the same Bacteroides-specific minimal media, this time with individual HMOs, glucose or lactose as carbon sources (0.5% w/v). All experiments were conducted with two biological replicates and three technical replicates per carbon source, under anaerobic conditions. Growth (OD600) was monitored every 30 minutes for 60h, at 37°C, using the Epoch2 plate reader (Agilent).
Optical density data over time was analyzed using an in-house custom script, the pheatmap (RRID : SCR_016418) and the growthcurver (Sprouffske and Wagner, 2016) packages under R statistical language [R Core Team (2021)].
RNA Sequencing of B. dorei Cultures
Pure cultures of B. dorei growing on various carbon sources (in replicates) were harvested at log phase (OD ~ 0.7) using the Direct-zol™ RNA Miniprep Plus kit (Zymo research, R2071). Agilent 2100 Bioanalyzer (Agilent Technologies) was employed for RNA quality control. Samples were processed according to a previously described protocol (Shishkin et al., 2015) and sequenced in two separate pools: HMOs (2’-FL, DFL, 3’-SL, 6’-SL, LNT, LNnT) and glucose. Single-end 75bp sequencing was performed on a NextSeq device.
The raw sequencing data were further filtered by trim_galore (https://github.com/FelixKrueger/TrimGalore), and classification of the B. dorei isolate to the strain level was achieved by alignment of reads using BLAST (Altschul et al., 1990). Principal Component Analysis (PCA) of transcriptional profiles on the various carbon sources was performed on the VST (variance stabilizing transformation)-transformed data (Love et al., 2014). Genome mapping to B. dorei DSM 17855 genome (GenBank: CP046176.1) was performed by Bowtie2 (Langmead and Salzberg, 2012), and differential expression analysis between HMOs and glucose was carried out using featureCounts (Liao et al., 2014) and DEseq2 (Love et al., 2014). Significantly up-regulated genes were selected based on the padj < 0.05 and log2 fold-change > 1 parameters. Gene annotation of glycoside hydrolases (GHs) was downloaded from the CAZy (Carbohydrate Active enZYmes) (Lombard et al., 2014) database. The statistical enrichment of GH genes from multiple families among the up-regulated gene group, compared to all B. dorei’s genes, was calculated with hypergeometric test, using the phyper function (stats package, R version 4.1.1).
Results
An Optimized System for Isolation of Bacteroides Strains From Infant Stool
To experimentally test the ability of infant Bacteroides strains to utilize HMOs as a carbon source, we first need to be able to isolate these infant strains from infant stool samples. In general, the ease of strain isolation differs depending on the species, however, infant stool samples may add an additional layer of complexity due to the low bacterial biomass (compared to adult stool) (Mitchell et al., 2020). Since the isolation process is laborious and time consuming, we aimed to apply it on a selective subset of samples for which we are confident that they harbor Bacteroides strains. The earliest possible step to identify the samples containing Bacteroides strains is following DNA extraction, for example, by using qPCR. We found that the previously published g-Bfra-F/R (Matsuki et al., 2002) primers enabled a clear distinction between Bacteroides-containing and non-Bacteroides-containing samples (Figure 1A). To control for differences in general bacterial DNA content, we compared the Bacteroides-specific Ct value to the universal bacterial 16S Ct value (Figure 1A).
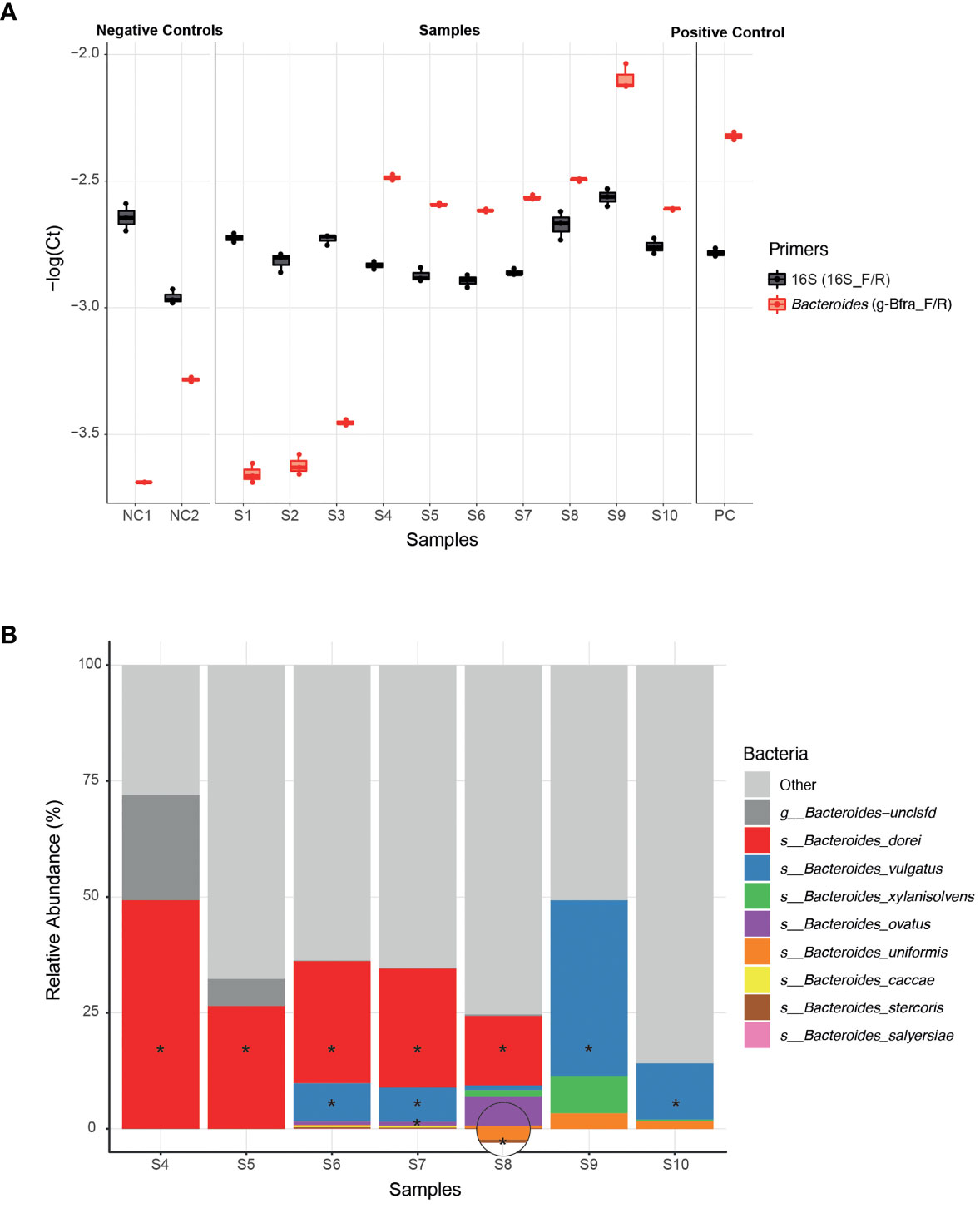
Figure 1 Isolation of Bacteroides strains from infant stool samples. (A) A comparison of qPCR Ct (Cycle threshold; Methods) values of infant stool samples DNA, using Bacteroides-specific primers (g-Bfra-F, g-Bfra-R) and universal 16S primers (16_F, 16_R). Each qPCR reaction was performed in triplicates, and each replicate is indicated by a dot. (B) Relative abundance of Bacteroides species, as identified by 16S rRNA gene sequencing of stool samples found to contain Bacteroides according to qPCR screening. Bacteroides species that were successfully isolated from stool samples are indicated with asterisks.
Next, we turned to optimize a system for isolation of Bacteroides species from samples with probable Bacteroides strains. We found that plating stool samples on a rich medium (such as blood agar plates) prior to plating on a more “harsh” selective medium, resulted in enhanced growth of Bacteroides and higher isolation success rates, perhaps due to the challenging nature of these Bacteroides strains (Medeiros, 1972; Pricop et al., 2020). Thus, we first plated the samples on blood agar plates, then we re-streaked multiple colonies on Bile Esculin Agar (BEA) Bacteroides-specific plates (Livingston et al., 1978). BEA plates contain both gentamicin, for which Bacteroides are inherently resistant to, and esculin, which Bacteroides are able to hydrolyze, producing a black pigment. Then, we picked single black colonies, and performed a second qPCR with Bacteroides-specific primers to exclude any non-Bacteroides gentamicin-resistant esculin hydrolyzing bacteria that might be present in the stool sample. Finally, Sanger sequencing of the 16S region was performed in order to receive an identification of the bacteria isolated at the genus level.
Indeed, we were able to successfully isolate 11 Bacteroides strains from infant samples that passed our qPCR filtering step (S4-S10, Figure 1A), whereas no Bacteroides were detected after plating and 16S sequencing of the samples that failed the qPCR step (S1-S3, Figure 1A).In addition, to test that the isolated strains match the overall Bacteroides composition of each sample, we also performed 16S sequencing the samples S4-S10 (Figure 1B and Supplementary Table 1). All isolated strains belong to species that were identified in the 16S sequencing of each sample. We were able to isolate strains at least from the most abundant species in each sample, and sometimes even from less abundant species, indicating the efficiency of our isolation method (Figure 1B).
Variation in HMO Utilization Across Bacteroides Isolates
Once we have established a strain collection composed of all successfully isolated Bacteroides strains from infant stool, we next wanted to examine the growth of the unique strains on different complex carbon sources. While there are many types of HMOs, in our study we use synthetic versions of HMOs from all three sub-types, including neutral, fucosylated and sialylated sugars: Lacto-N-Tetraose (LNT), Lacto-N-neotetraose (LNnT), 2′-fucosyllactose (2′-FL), difucosyllactose (DFL), 3-Sialyllactose (3’-SL), and 6-Sialyllactose (6’SL). Growth curve assays on some of the isolated strains were performed with Bacteroides-specific minimal media (MM) mixed with the chosen carbon source to a final concentration of 0.5% (weight/volume). Our results indicate that our isolated Bacteroides strains differ in their HMO utilization capability and dynamics (Figures 2A, B and Supplementary Figure 1). Some strains do not grow on any tested HMO as a single carbon source (B. stercoris), whereas others exhibit growth on various sugars (B. doeri 2, B. vulgatus; Figures 2A, B and Supplementary Figure 1). Furthermore, we also observed intra-species variation, in both B. vulgatus and B. dorei strains, where two strains of the same species exhibit distinct utilization patterns. As one might expect, although it has not been reported yet, strains tended to display similar growth characteristics on similarly structured HMOs, for example, the growth pattern of B. dorei 2 on 3’SL is similar to its growth on 6’SL, and these sugars are both sialylated (Figure 2B).
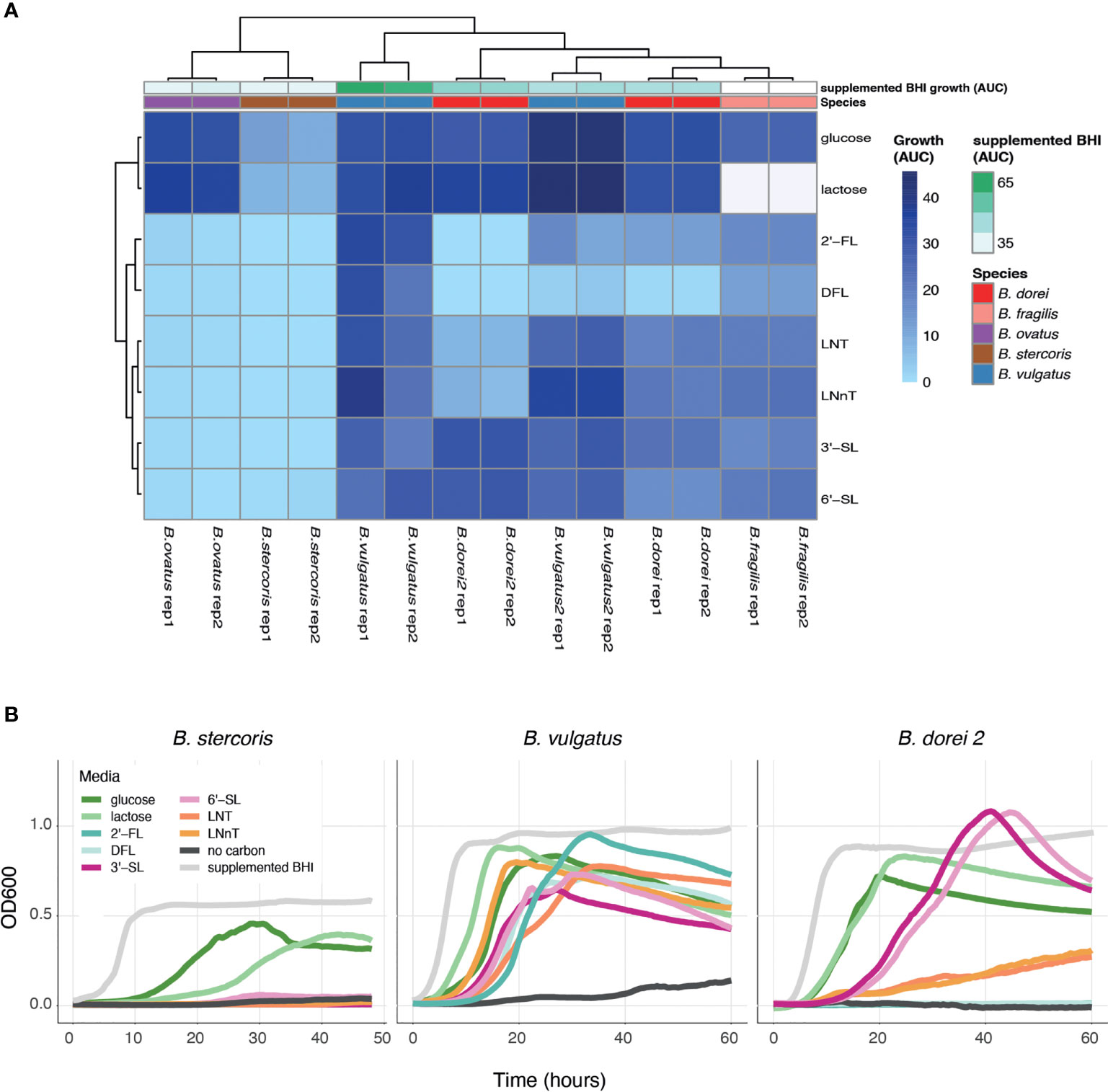
Figure 2 HMO utilization ability varies across Bacteroides isolates. Growth curves were measured for all chosen strains and all chosen sources (0.5% weight/volume): glucose, lactose, 2-Fucosyllactose (2’-FL), Difucosyllactose (DFL), 3-Sialyllactose (3’-SL), 6-Sialyllactose (6’-SL), Lacto-N-Tetraose (LNT) and Lacto-N-neotetraose (LNnT), supplemented BHI media (positive control), no carbon media (negative control). (A) Area under the curve (AUC) of the growth curve plots, shown in two biological replicates for each strain. Missing data is represented by white cells, in both heat map and annotations. (B) Growth curve plots of B. stercoris, B. vulgatus and B. dorei 2 on all selected carbon sources.
To make sure that the strains cannot grow on any of the minimal media (MM) components on its own, the negative control in our growth experiments was MM without an added carbon source. The positive control was supplemented BHI media, which is a rich nonselective media that supports the growth of many bacteria. Some strains are inherently more difficult to grow in the lab, and thus we used the positive control to evaluate the ease of growth for each strain. In addition, we also measure the strains’ ability to grow on glucose and lactose (the main carbohydrate in breast-milk) (Ballard and Morrow, 2013). Growth experiments can be evaluated by the kinetics of growth (using the OD600 over time), and by emphasizing the quantitative ability of a strain to utilize a carbon source (using the area under the OD600 curve; AUC; Figure 2A). The AUC statistic was calculated for all experiments, varied across carbon sources, yet showed consistency across biological replicates (Figure 2A).
Searching for B. dorei’s Differentially Expressed Genes Across Multiple HMO Conditions
Differential expression of genes under various conditions can shed light on the enzymes, mechanisms and pathways enabling the utilization of the specific carbon sources. We next chose the isolate of Bacteroides dorei that was able to utilize all HMOs tested to some extent, B. dorei 1 (Supplementary Figure 1), to search the genes that enable this utilization. We want to compare the transcribed genes in each condition to identify the upregulated genes that can account for the HMO utilization capability. We grew B. dorei on the full set of HMOs described above and harvested the cultures during the mid-log phase to perform transcriptional profiling using RNA sequencing (RNA-Seq).
To identify the genes up regulated by B. dorei during HMO consumption, whole-genome transcriptional profiling of B. dorei on MM supplemented with individual HMOs was compared to growth on MM-glucose (Supplementary Table 2 and Supplementary Figure 2). The two biological replicates of each condition were similar to one another, and more similar within a condition than across conditions (Supplementary Figure 3). Fucosylated sugars (2’-FL, DFL) clustered separately from sialylated (3’-SL, 6’-SL), and HMO core structures (LNT, LNnT) clustered in-between (Figure 3A). A possible explanation could be that LNT and LNnT lack unique sialyl or fucosyl residues distinguishing them from the other sugars. Overall, the examined conditions are relatively similar, as the tested sugars differ from one another by small modifications only. Thus, we expected the overall gene-expression profiles to be rather similar, with small, yet distinct differences, across all conditions (Supplementary Figure 3).
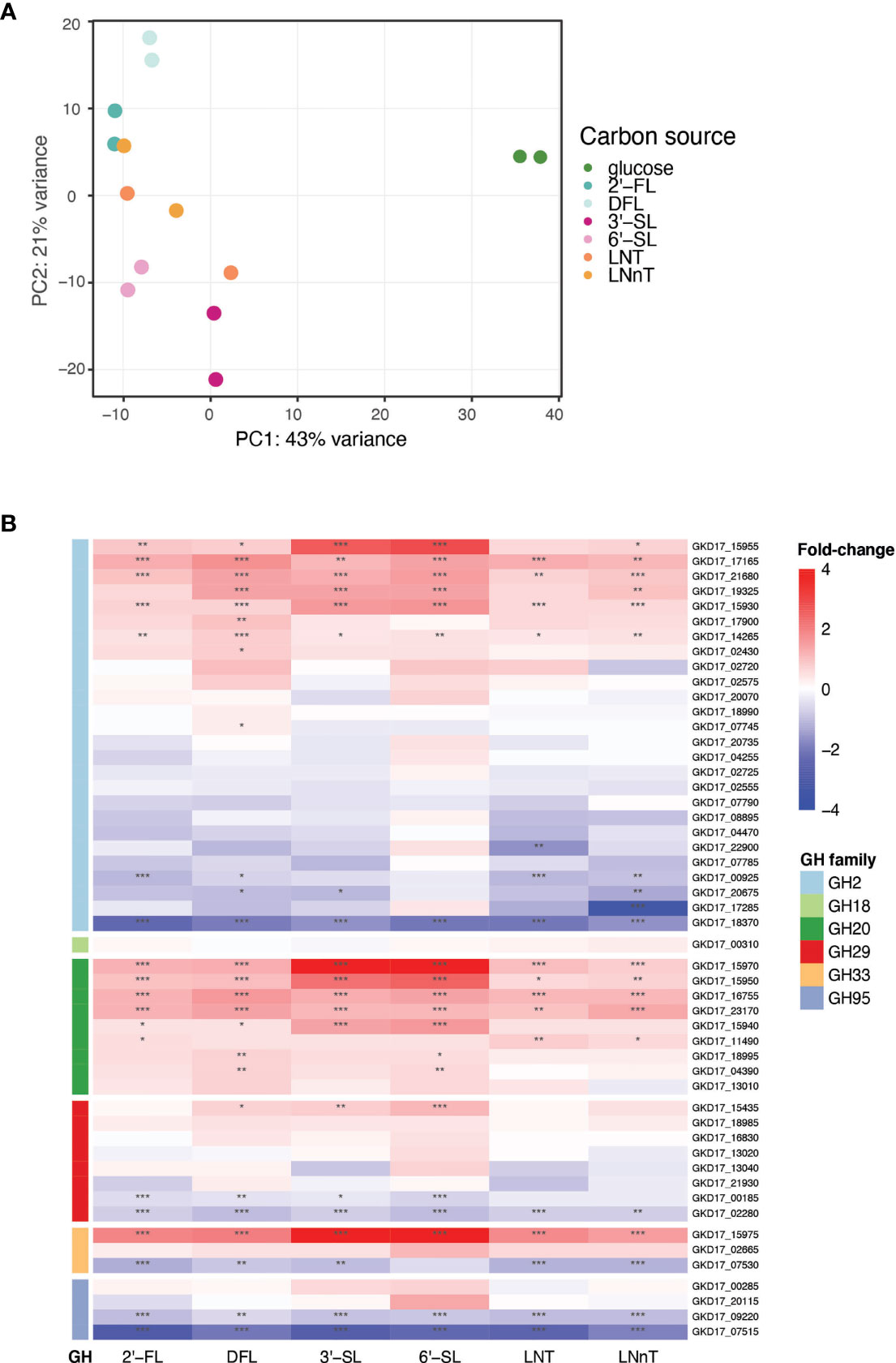
Figure 3 B. dorei’s transcriptional response to HMOs is similar across conditions. (A) Principal Component Analysis (PCA) of transcriptional profiles of pure cultures of B. dorei on various carbon sources, in two biological replicates, using RNA-Sequencing (Methods). (B) Heat map of log2 fold-change of genes (rows) belonging to glycoside hydrolase (GH) families previously known to break down HMOs. Gene expression data on HMOs (2’-FL, DFL, 3’-SL, 6’-SL, LNT, LNnT; columns) was compared to gene expression on glucose. p < 0.05 (*); p < 0.01 (**); p < 0.001 (***).
B. dorei possesses a repertoire of predicted glycoside hydrolases (GHs), carbohydrate active enzymes (CAZy) capable of breaking the carbon bond found in all human milk oligosaccharides. B. dorei’s 213 glycoside hydrolases comprise 59 unique GH families (Lombard et al., 2014). Differential expression analysis of the RNA-Seq data revealed that B. dorei exhibits an expansive glycoside hydrolase response during consumption of HMOs in vitro. Many unique GH families were upregulated (log2foldChange > 1, padj < 0.05) when growing on HMOs compared to glucose: 17 GH families were upregulated when grown on 2’-FL, 21 on DFL, 19 on 3’-SL, 23 on 6’-SL, 15 on LNT, and 18 on LNnT.
First, we examined the differential expression of GH families previously annotated as capable of breaking down the HMO carbon bond (Marcobal et al., 2011): GH2, GH18, GH20, GH29, GH33, and GH95 (Figure 3B). Two additional GH families have been reported in the literature (GH85, GH112) (Ioannou et al., 2021), however they are not encoded in the B. dorei genome. Most research regarding HMO utilization by infant gut microbes has focused on Bifidobacterium species, hence the knowledge on Bacteroides cellular response to HMOs is rather limited. Indeed, some GH families that are well annotated in Bifidobacterium species did not exhibit any clear differential expression here, like GH18 and GH95. Surprisingly, some of the GH95 and GH29 B. dorei genes, which break the α1-2 and α1-3/4 fucosyl bonds, accordingly, were up-regulated in non-fucosylated HMOs. This lack of specificity is unexpected as we would expect GH95 and GH29 to be upregulated only on fucosylated HMOs. Similarly, GH33 which is responsible for sialic acid release from sialylated HMOs, was up-regulated across all HMO conditions (sialylated and non-sialylated HMOs), although the up-regulation in sialylated oligosaccharides is the most prominent. Lastly, in GH2, the largest GH family in B. dorei’s genome (Lombard et al., 2014), some genes were up-regulated while others were down regulated, which might be confusing, and is further discussed below.
Multiple Glycoside Hydrolase Families Are Upregulated During B. dorei’s Growth on HMOs
We next performed an unbiased search for all up-regulated GH families in our data, regardless of their annotations. To identify the GH families that could be responsible for B. dorei’s HMO utilization, we searched for all GH families that were upregulated in similarly-structured HMO-pairs: 2’-FL and DFL, 3’-SL and 6’-SL, LNT and LNnT (Figure 4A). A GH family that is upregulated in two structurally-similar HMOs is more likely to play a role in breaking down these common carbon bonds.
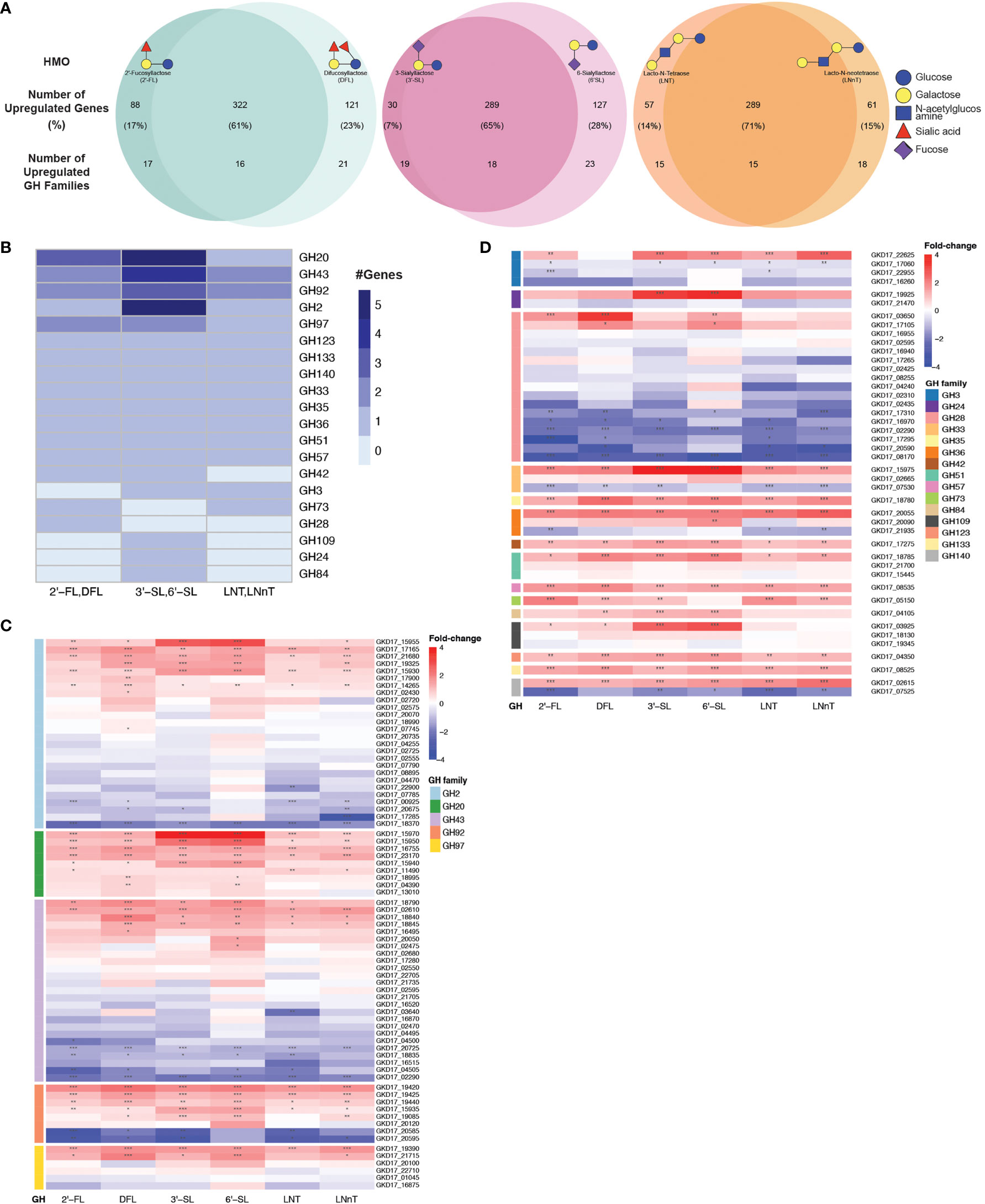
Figure 4 B. dorei shares up-regulated genes upon growth on HMOs compared to glucose. (A) Venn diagrams of B. dorei’s up-regulated genes and glycoside hydrolase (GH) families across HMO types, highlighting the similar transcriptional response to HMOs, regardless of specific conditions. HMOs were divided into pairs based on structural similarity. Percentage values refer to the fraction of up-regulated genes from the total up-regulated genes in both members of an HMO pair. (B) A heat map showing the number of up-regulated genes in each up-regulated GH family in at least one HMO pair. (C, D) Heat maps of log2 fold-change of genes (rows) belonging to GH families with (C) at least two; or (D) a single up-regulated gene(s). Gene expression data on HMOs (2’-FL, DFL, 3’-SL, 6’-SL, LNT, LNnT; columns) was compared to gene expression on glucose. p < 0.05 (*); p < 0.01 (**); p < 0.001 (***).
图 4 与葡萄糖相比,B. dorei 在 HMO 上生长时共享上调的基因。(A) B. dorei 上调基因和不同 HMO 类型的糖苷水解酶 (GH) 家族的维恩图,突出了对 HMO 的相似转录反应,无论具体条件如何。根据结构相似性将 HMO 分为两对。百分比值是指 HMO 对的两个成员中上调基因占上调基因总数的比例。(B) 热图显示至少一对 HMO 中每个上调的 GH 家族中上调的基因数量。(C、D)属于 GH 家族的基因(行)的 log2 倍变化热图 (C) 至少有两个;或 (D) 单个上调基因。将 HMO(2'-FL、DFL、3'-SL、6'-SL、LNT、LNnT;列)的基因表达数据与葡萄糖的基因表达数据进行比较。p < 0.05 (*);p < 0.01 (**);p < 0.001 (***)。
First, we focused on GH families that had a minimum of two upregulated genes for at least one HMO-pair, and found five such GH families: GH2, G20, GH43, GH92, and GH97 (Figure 4B). We then expanded our analysis to all genes that belong to these families to compare their regulation across the various HMO structures (Figure 4C). We did not find a specific GH family response for a specific HMO-pair, rather, it seems that B. dorei’s response to growth on various HMOs is quite generalized, regardless of the specific HMO molecule. Furthermore, even though the response is not specific towards a single HMO, it is specific in the transcriptional changes. Namely, in most cases, only a small subset of the genes in each GH family were up-regulated, and the others were often even down-regulated, suggesting that a specific set of genes, rather than all genes in a certain GH family, responded to the change in carbon source. For example, we found that among B. dorei’s 27 GH2 genes, two genes only (GKD17_17165, GKD17_15930) were consistently, significantly upregulated for all conditions. Likewise, for GH43 and GH97, GKD17_02610 and GKD17_19390 were the only consistently upregulated genes, respectively (Figure 4C).
Second, we were interested to inspect if the pattern of simultaneous up- and down-regulation of different genes from the same GH family is evident for additional GH families. We looked at GH families for which only a single gene was upregulated in at least one HMO-pair (Figure 4D), as a single up-regulated gene can be sufficient for HMO utilization. Similarly to the analysis in the first step (Figure 4C), oftentimes, for each GH family, only one or two specific genes were consistently up-regulated, while other members of the family were down-regulated compared to MM-glucose. This observation of specificity in upregulated genes was also evident in smaller GH families, such as GH3 (GKD17_22625), GH33 (GKD17_15975), GH36 (GKD17_20055), GH51 (GKD17_18785). Lastly, GH140 with only two genes presented an interesting pattern, where one was strongly upregulated (GKD17_02615) while the other was strongly down-regulated (GKD17_07525), which may merit further study of this particular GH family. Interestingly, when looking at all GH28 genes encoded in B. dorei’s genome, our results suggest that all conditions invoke a strong down-regulation response for many of the GH genes. This consistent broad down-regulation might indicate the up-regulation of other, yet to be annotated, GH genes.
Additionally, enrichment analysis of the shared upregulated GH families revealed that some GH families were enriched in the upregulated gene group compared to the total amount of genes in B. dorei’s genome: GH20, GH43 and GH92 were significantly (p<0.05) enriched for DFL, 3’-SL and 6’-SL, GH3 was significantly (p<0.05) enriched for LNT, LNnT, 3’-SL and 6’-SL, GH2 was significantly (p<0.05) enriched for 3’-SL and 6’-SL, and GH36 was significantly (p<0.05) enriched for 6’-SL.
Discussion
In our quest to investigate HMO utilization by Bacteroides infant strains, we have established a system for screening, culturing, measuring growth curves and performing RNA sequencing of Bacteroides isolates. We constructed growth curves of Bacteroides isolates from infant stool and demonstrated variation in HMO-utilization patterns across Bacteroides species and within strains of the same species. Some Bacteroides isolates do not grow on HMOs at all, whereas others can utilize all tested HMOs to some extent. This is consistent with previous studies, showing that Bacteroides are metabolically diverse and are capable of utilizing many versatile carbon sources (Salyers et al., 1977; Sonnenburg et al., 2005; Bjursell et al., 2006; Marcobal et al., 2011; Wexler and Goodman, 2017; Carrow et al., 2020; García-Bayona and Comstock, 2020; Fultz et al., 2021). Pioneering work in Bacteroides demonstrated B. thetaiotomicron’s ability to expand and adapt its metabolism, from mainly utilizing glucose and lactose, to also being able to break down more complex plant polysaccharides (Bjursell et al., 2006). Thus, species isolated from infant stool at different time points might alter their metabolic repertoire as the infant ages. A strain’s ability to utilize a wide variety of HMOs may confer a selective advantage over strains that do not utilize these sugars, and could select for its survival in the infant gut.
Our growth curve results demonstrate similar growth patterns on similarly structured HMOs, which may suggest unique carbohydrate utilization pathways being activated for HMOs with similar residues. Bacteroides are known to have a wide range of polysaccharide utilization loci (PUL) that encode common classes of proteins involved in polysaccharide utilization, and a large percentage of their genome is dedicated to the sensing, import, and hydrolysis of diverse glycans (Bjursell et al., 2006; Martens et al., 2008). However, RNA-Seq experiments do not show unique genes upregulated under specific conditions. When examining differential expression of glycoside hydrolase (GH) genes of various families, a generalized HMO response was observed, with several GH families upregulated across all six HMOs tested: 2’-FL, DFL, 3’-SL, 6’-SL, LNT, and LNnT. Hence, our findings suggest a unique HMO-utilization strategy among some Bacteroides strains, diverging from the extensively studied Bifidobacterium HMO utilization mechanisms, where distinct enzymes were recognized for specific carbon bonds in HMO molecules. A possible explanation for the similar response of B. dorei to all tested HMOs could be that overall, the conditions tested are rather close, with identical minimal media composition for all samples and small changes in HMO modifications.
Bifidobacterium species are considered to be well-adapted to HMO utilization, and they have evolved two distinct strategies for this purpose. One is transporter-dependent (intracellular digestion strategy), and the other is extracellular glycosidase-dependent (extracellular digestion strategy), utilizing the same enzymes, only in a membrane-bound form (Sela et al., 2008; Turroni et al., 2014; Bunesova et al., 2016). Both strategies employ specific enzymes (Ioannou et al., 2021), some of them clustered in specific genomic regions (Sela et al., 2008; LoCascio et al., 2010) whereas others are spread across additional genomic locations (Zabel et al., 2020). In contrast, very little is known about Bacteroides HMO utilization, and only a few papers have discussed this issue in terms of phenotypic growth (Salli et al., 2021) or transcriptional profiling (Marcobal et al., 2011). Due to the high-prevalence of Bacteroides in the infant gut and a seeming deviation from the traditional Bifidobacterium-dominated infant gut microbiome (Vatanen et al., 2019; Casaburi et al., 2021), it is increasingly important to decipher the HMO utilization strategies employed by Bacteroides species as well.
B. dorei’s differential expression pattern in response to HMOs does not appear to be HMO-specific, however, we show that for the upregulated GH families in our data, a particular set of genes is up-regulated among all members of the GH family encoded in the genome. Moreover, the remaining members of the GH families explored are down-regulated, perhaps meaning that B. dorei’s response to HMOs is tailored to exploit some GH genes over others. The novel pattern of HMO utilization presented here can indicate that for a given Bacteroides strain, we can discuss specific genes responsible for HMO break-down, in addition to discussing the results on the broader, less-selective, GH family level.
The pioneering work of exploring HMO utilization mechanisms by the type strains of B. thetaiotaomicron and B. fragilis (Marcobal et al., 2011) found that certain genes are heavily upregulated upon growth on HMOs. These genes belong to several GH families: GH2, GH18, GH20, GH29, GH33, and GH95. There are a few notable differences between this early study and our results. First, the researchers used an HMO mixture, while we used individual HMO molecules. Second, to our knowledge, no study has looked at the transcriptional response of Bacteroides strains that were isolated from infant sool to HMOs, which could be different from the type strain response, due to large variation across Bacteroides strains of the same species. Lastly, in the 2011 manuscript, the transcriptional response was examined on a pre-selected set of genes (using microarrays designed to capture these specific RNA molecules), whereas we used an unbiased approach of total RNA sequencing. These differences make it difficult to compare the results of both studies, highlighting the need for additional research in additional species, preferably in an unbiased manner.
Notably, the Bacteroides genus is characterized by great variations in carbon-utilization abilities, both across species and within strains of the same species (Lange et al., 2016; Husain et al., 2017; Pasolli et al., 2019). Therefore, our findings, exploring one isolate only, do represent a general HMO utilization pattern, and every Bacteroides strain or isolate of interest should be individually examined. Second, we compared gene expression profiles of B. dorei cultures grown with various HMOs as a single carbon source, in comparison to MM-glucose only. While glucose is the most common reference in microbiology literature, follow up studies should also include galactose and lactose as a reference. Lastly, we should keep in mind that bacteria in the infant gut are simultaneously exposed to multiple types of HMOs, in the context of a complex microbial ecosystem. Experiments such as the ones presented test the utilization capacity of individual strains with a specific HMO molecule. These studies are critical stepping stones towards constructing a comprehensive map of how the infant gut microbes can utilize glycans found in breastmilk, thus shedding light on how breastfeeding, and breastmilk composition, impact the nursing infant gut microbiome.
Data Availability Statement
The data presented in the study are deposited in the National Center for Biotechnology Information (NCBI) repository, accession number PRJNA804725.
Ethics Statement
The studies involving human participants were reviewed and approved by Hebrew University’s Institutional Review Board (IRB) (approval number 20042021). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
SK - Established the experimental system, performed experiments and analysis, and wrote the manuscript. AC- Assisted in establishing the experimental system. MY- Guided the work and wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was funded in part by the Azrieli Foundation grant for faculty fellows. SK, AC, and MY were supported by the Azrieli Foundation and MY was also supported by the Israel Science Foundation grant 2660/18.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2022.854122/full#supplementary-material
Supplementary Figure 1 | (A) Growth curves of B. dorei, B. vulgatus 2, B. ovatus, and B. fragilis on various HMOs (2’-FL, DFL, 3’-SL, 6’-SL, LNT and LNnT), glucose, lactose, supplemented BHI media (positive control), and no carbon media (negative control). (B) Principal Component Analysis (PCA) of Bacteroides isolates based on the ability to grow on all media types, as measured by the area under the curve (AUC) of each growth curve.
Supplementary Figure 2 | MA plots of HMO transcriptional profiles compared to glucose, shown as M (log ratio) on the y-axis and A (mean average) on the x-axis. Here, the log ratio is calculated as the log2 of the change in expression values of the HMOs vs. glucose, and the mean average is of the glucose read counts. Genes from GH families are labeled with larger dots and display the GH family they belong to. The plot highlights up-regulated (red) and down-regulated (blue) genes, using a threshold of 1 for the absolute log2 fold change, and a threshold of 4 for the log2 glucose mean read count.
Supplementary Figure 3 | B. dorei’s transcriptional profile, as characterized by RNA-Seq experiments, is similar across conditions. The scatterplots display the normalized read count value for all annotated genes in B. dorei’s genome (dots), compared between replicates of the same carbon source (diagonal plots) and between different carbon sources (bottom triangle). Pearson coefficient was calculated for each comparison.
Supplementary Table 1 | Bacterial relative abundances (%) as detected by 16S ribosomal RNA gene sequencing, at the species level.
Supplementary Table 2 | B. dorei’s transcriptional response upon growth in minimal media (MM) supplemented with various HMOs, compared to MM-glucose. Each sheet represents a different HMO compared to glucose, and the last sheet contains explanations of all columns in previous sheets.
References
Al-Ghalith, G., Knights, D. (2020). BURST Enables Mathematically Optimal Short-Read Alignment for Big Data. Zenodo. doi: 10.1101/2020.09.08.287128. 2020.09.08.287128.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., Lipman, D. J. (1990). Basic Local Alignment Search Tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Aronesty, E. (2013). Comparison of Sequencing Utility Programs. Open Bioinforma. J. 7, 1–8. doi: 10.2174/1875036201307010001
Azad, M. B., Konya, T., Maughan, H., Guttman, D. S., Field, C. J., Chari, R. S., et al. (2013). Gut Microbiota of Healthy Canadian Infants: Profiles by Mode of Delivery and Infant Diet at 4 Months. CMAJ 185, 385–394. doi: 10.1503/cmaj.121189
Bäckhed, F., Roswall, J., Peng, Y., Feng, Q., Jia, H., Kovatcheva-Datchary, P., et al. (2015). Dynamics and Stabilization of the Human Gut Microbiome During the First Year of Life. Cell Host Microbe 17, 690–703. doi: 10.1016/j.chom.2015.04.004
Bai, Y., Tao, J., Zhou, J., Fan, Q., Liu, M., Hu, Y., et al. (2018). Fucosylated Human Milk Oligosaccharides and N-Glycans in the Milk of Chinese Mothers Regulate the Gut Microbiome of Their Breast-Fed Infants During Different Lactation Stages. mSystems 3, e00206–18. doi: 10.1128/mSystems.00206-18
Ballard, O., Morrow, A. L. (2013). Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. North Am. 60, 49–74. doi: 10.1016/j.pcl.2012.10.002
Bjursell, M. K., Martens, E. C., Gordon, J. I. (2006). Functional Genomic and Metabolic Studies of the Adaptations of a Prominent Adult Human Gut Symbiont, Bacteroides Thetaiotaomicron, to the Suckling Period. J. Biol. Chem. 281, 36269–36279. doi: 10.1074/jbc.M606509200
Borewicz, K., Gu, F., Saccenti, E., Arts, I. C. W. (2019). Correlating Infant Fecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of 1-Month-Old Breastfed Infants. Mol. Nutr. Food Res. 63 (13), e1801214. doi: 10.1002/mnfr.201801214
Borewicz, K., Gu, F., Saccenti, E., Hechler, C., Beijers, R., de Weerth, C., et al. (2020). The Association Between Breastmilk Oligosaccharides and Faecal Microbiota in Healthy Breastfed Infants at Two, Six, and Twelve Weeks of Age. Sci. Rep. 10, 4270. doi: 10.1038/s41598-020-61024-z
Buffie, C. G., Pamer, E. G. (2013). Microbiota-Mediated Colonization Resistance Against Intestinal Pathogens. Nat. Rev. Immunol. 13, 790–801. doi: 10.1038/nri3535
Bunesova, V., Lacroix, C., Schwab, C. (2016). Fucosyllactose and L-Fucose Utilization of Infant Bifidobacterium Longum and Bifidobacterium Kashiwanohense. BMC Microbiol. 16, 248. doi: 10.1186/s12866-016-0867-4
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Huntley, J., Fierer, N., et al. (2012). Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. ISME J. 6, 1621–1624. doi: 10.1038/ismej.2012.8
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global Patterns of 16S rRNA Diversity at a Depth of Millions of Sequences Per Sample. Proc. Natl. Acad. Sci. U. S. A. 108 Suppl 1, 4516–4522. doi: 10.1073/pnas.1000080107
Carrow, H. C., Batachari, L. E., Chu, H. (2020). Strain Diversity in the Microbiome: Lessons From Bacteroides Fragilis. PloS Pathog. 16, e1009056. doi: 10.1371/journal.ppat.1009056
Casaburi, G., Duar, R. M., Brown, H., Mitchell, R. D., Kazi, S., Chew, S., et al. (2021). Metagenomic Insights of the Infant Microbiome Community Structure and Function Across Multiple Sites in the United States. Sci. Rep. 11, 1472. doi: 10.1038/s41598-020-80583-9
Chichlowski, M., Shah, N., Wampler, J. L., Wu, S. S., Vanderhoof, J. A. (2020). Bifidobacterium Longum Subspecies Infantis (B. Infantis) in Pediatric Nutrition: Current State of Knowledge. Nutrients 12, 1581–1599. doi: 10.3390/nu12061581
Davis, J. C. C., Totten, S. M., Huang, J. O., Nagshbandi, S., Kirmiz, N., Garrido, D. A., et al. (2016). Identification of Oligosaccharides in Feces of Breast-Fed Infants and Their Correlation With the Gut Microbial Community. Mol. Cell. Proteomics 15, 2987–3002. doi: 10.1074/mcp.M116.060665
Dethlefsen, L., McFall-Ngai, M., Relman, D. A. (2007). An Ecological and Evolutionary Perspective on Human-Microbe Mutualism and Disease. Nature 449, 811–818. doi: 10.1038/nature06245
Dominguez-Bello, M. G., Costello, E. K., Contreras, M., Magris, M., Hidalgo, G., Fierer, N., et al. (2010). Delivery Mode Shapes the Acquisition and Structure of the Initial Microbiota Across Multiple Body Habitats in Newborns. Proc. Natl. Acad. Sci. U. S. A. 107, 11971–11975. doi: 10.1073/pnas.1002601107
Engfer, M. B., Stahl, B., Finke, B., Sawatzki, G., Daniel, H. (2000). Human Milk Oligosaccharides are Resistant to Enzymatic Hydrolysis in the Upper Gastrointestinal Tract. Am. J. Clin. Nutr. 71, 1589–1596. doi: 10.1093/ajcn/71.6.1589
Favier, C. F., Vaughan, E. E., De Vos, W. M., Akkermans, A. D. L. (2002). Molecular Monitoring of Succession of Bacterial Communities in Human Neonates. Appl. Environ. Microbiol. 68, 219–226. doi: 10.1128/AEM.68.1.219-226.2002
Ferretti, P., Pasolli, E., Tett, A., Asnicar, F., Gorfer, V., Fedi, S., et al. (2018). Mother-To-Infant Microbial Transmission From Different Body Sites Shapes the Developing Infant Gut Microbiome. Cell Host Microbe 24, 133–145.e5. doi: 10.1016/j.chom.2018.06.005
Fultz, R., Ticer, T., Ihekweazu, F. D., Horvath, T. D., Haidacher, S. J., Hoch, K. M., et al. (2021). Unraveling the Metabolic Requirements of the Gut Commensal Bacteroides Ovatus. Front. Microbiol. 12, 745469. doi: 10.3389/fmicb.2021.745469
García-Bayona, L., Comstock, L. E. (2020). Utilizing Ribose Compounds: How Bacteroides PUL It Off. Cell Host Microbe 27, 6–8. doi: 10.1016/j.chom.2019.12.010
Gnoth, M. J., Kunz, C., Kinne-Saffran, E., Rudloff, S. (2000). Human Milk Oligosaccharides are Minimally Digested In Vitro. J. Nutr. 130, 3014–3020. doi: 10.1093/jn/130.12.3014
Harmsen, H. J., Wildeboer-Veloo, A. C., Raangs, G. C., Wagendorp, A. A., Klijn, N., Bindels, J. G., et al. (2000). Analysis of Intestinal Flora Development in Breast-Fed and Formula-Fed Infants by Using Molecular Identification and Detection Methods. J. Pediatr. Gastroenterol. Nutr. 30, 61–67. doi: 10.1097/00005176-200001000-00019
Hegar, B., Wibowo, Y., Basrowi, R. W., Ranuh, R. G., Sudarmo, S. M., Munasir, Z., et al. (2019). The Role of Two Human Milk Oligosaccharides, 2′-Fucosyllactose and Lacto-N-Neotetraose, in Infant Nutrition. Pediatr. Gastroenterol. Hepatol. Nutr. 22, 330–340. doi: 10.5223/pghn.2019.22.4.330
Hooper, L. V., Midtvedt, T., Gordon, J. I. (2002). How Host-Microbial Interactions Shape the Nutrient Environment of the Mammalian Intestine. Annu. Rev. Nutr. 22, 283–307. doi: 10.1146/annurev.nutr.22.011602.092259
Husain, F., Tang, K., Veeranagouda, Y., Boente, R., Patrick, S., Blakely, G., et al. (2017). Novel Large-Scale Chromosomal Transfer in Bacteroides Fragilis Contributes to its Pan-Genome and Rapid Environmental Adaptation. Microb. Genom. 3, e000136. doi: 10.1099/mgen.0.000136
Ioannou, A., Knol, J., Belzer, C. (2021). Microbial Glycoside Hydrolases in the First Year of Life: An Analysis Review on Their Presence and Importance in Infant Gut. Front. Microbiol. 12, 631282. doi: 10.3389/fmicb.2021.631282
Ivanov, I. I., Atarashi, K., Manel, N., Brodie, E. L., Shima, T., Karaoz, U., et al. (2009). Induction of Intestinal Th17 Cells by Segmented Filamentous Bacteria. Cell 139, 485–498. doi: 10.1016/j.cell.2009.09.033
Jacobi, S. K., Yatsunenko, T., Li, D., Dasgupta, S., Yu, R. K., Berg, B. M., et al. (2016). Dietary Isomers of Sialyllactose Increase Ganglioside Sialic Acid Concentrations in the Corpus Callosum and Cerebellum and Modulate the Colonic Microbiota of Formula-Fed Piglets. J. Nutr. 146, 200–208. doi: 10.3945/jn.115.220152
Lane, D. J. (1991). 16S/23S rRNA Sequencing. In: Stackebrandt, E., Goodfellow, M., Eds., Nucleic Acid Techniques in Bacterial Systematic. (New York: John Wiley and Sons), 115–175. doi: 10.1177/016344391013001010
Lange, A., Beier, S., Steimle, A., Autenrieth, I. B., Huson, D. H., Frick, J.-S. (2016). Extensive Mobilome-Driven Genome Diversification in Mouse Gut-Associated Bacteroides Vulgatus Mpk. Genome Biol. Evol. 8, 1197–1207. doi: 10.1093/gbe/evw070
Langmead, B., Salzberg, S. L. (2012). Fast Gapped-Read Alignment With Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
La Rosa, P. S., Warner, B. B., Zhou, Y., Weinstock, G. M., Sodergren, E., Hall-Moore, C. M., et al. (2014). Patterned Progression of Bacterial Populations in the Premature Infant Gut. Proc. Natl. Acad. Sci. U. S. A. 111, 12522–12527. doi: 10.1073/pnas.1409497111
Laursen, M. F., Pekmez, C. T., Larsson, M. W., Lind, M. V., Yonemitsu, C., Larnkjær, A., et al. (2021). Maternal Milk Microbiota and Oligosaccharides Contribute to the Infant Gut Microbiota Assembly. ISME Commun. 1, 1–13. doi: 10.1038/s43705-021-00021-3
Lawson, M. A. E., O’Neill, I. J., Kujawska, M., Gowrinadh Javvadi, S., Wijeyesekera, A., Flegg, Z., et al. (2020). Breast Milk-Derived Human Milk Oligosaccharides Promote Bifidobacterium Interactions Within a Single Ecosystem. ISME J. 14, 635–648. doi: 10.1038/s41396-019-0553-2
Lax, S., Smith, D. P., Hampton-Marcell, J., Owens, S. M., Handley, K. M., Scott, N. M., et al. (2014). Longitudinal Analysis of Microbial Interaction Between Humans and the Indoor Environment. Science. 345, 1048–1052. doi: 10.1126/science.1254529
Letten, A. D., Baumgartner, M., Pfrunder-Cardozo, K. R., Levine, J. M., Hall, A. R. (2021a). Human-Associated Microbiota Suppress Invading Bacteria Even Under Disruption by Antibiotics. ISME J. 15, 2809–2812. doi: 10.1038/s41396-021-00929-7
Letten, A. D., Hall, A. R., Levine, J. M. (2021b). Using Ecological Coexistence Theory to Understand Antibiotic Resistance and Microbial Competition. Nat. Ecol. Evol. 5, 431–441. doi: 10.1038/s41559-020-01385-w
Liao, Y., Smyth, G. K., Shi, W. (2014). Featurecounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 30, 923–930. doi: 10.1093/bioinformatics/btt656
Liu, F., Yan, J., Wang, X., Wang, C., Chen, L., Li, Y., et al. (2021). Maternal Fucosyltransferase 2 Status Associates With the Profiles of Human Milk Oligosaccharides and the Fecal Microbiota Composition of Breastfed Infants. J. Agric. Food Chem. 69, 3032–3043. doi: 10.1021/acs.jafc.0c04575
Livingston, S. J., Kominos, S. D., Yee, R. B. (1978). New Medium for Selection and Presumptive Identification of the Bacteroides Fragilis Group. J. Clin. Microbiol. 7, 448–453. doi: 10.1128/jcm.7.5.448-453.1978
LoCascio, R. G., Desai, P., Sela, D. A., Weimer, B., Mills, D. A. (2010). Broad Conservation of Milk Utilization Genes in Bifidobacterium Longum Subsp. Infantis as Revealed by Comparative Genomic Hybridization. Appl. Environ. Microbiol. 76, 7373–7381. doi: 10.1128/AEM.00675-10
Lombard, V., Golaconda Ramulu, H., Drula, E., Coutinho, P. M., Henrissat, B. (2014). The Carbohydrate-Active Enzymes Database (CAZy) in 2013. Nucleic Acids Res. 42, D490–D495. doi: 10.1093/nar/gkt1178
Love, M. I., Huber, W., Anders, S. (2014). Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data With Deseq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
Madan, J. C., Hoen, A. G., Lundgren, S. N., Farzan, S. F., Cottingham, K. L., Morrison, H. G., et al. (2016). Association of Cesarean Delivery and Formula Supplementation With the Intestinal Microbiome of 6-Week-Old Infants. JAMA Pediatr. 170, 212–219. doi: 10.1001/jamapediatrics.2015.3732
Marcobal, A., Barboza, M., Sonnenburg, E. D., Pudlo, N., Martens, E. C., Desai, P., et al. (2011). Bacteroides in the Infant Gut Consume Milk Oligosaccharides via Mucus-Utilization Pathways. Cell Host Microbe 10, 507–514. doi: 10.1016/j.chom.2011.10.007
Marcobal, A., Sonnenburg, J. L. (2012). Human Milk Oligosaccharide Consumption by Intestinal Microbiota. Clin. Microbiol. Infect. 18 (Suppl 4), 12–15. doi: 10.1111/j.1469-0691.2012.03863.x
Martens, E. C., Chiang, H. C., Gordon, J. I. (2008). Mucosal Glycan Foraging Enhances Fitness and Transmission of a Saccharolytic Human Gut Bacterial Symbiont. Cell Host Microbe 4, 447–457. doi: 10.1016/j.chom.2008.09.007
Matsuki, T., Watanabe, K., Fujimoto, J., Miyamoto, Y., Takada, T., Matsumoto, K., et al. (2002). Development of 16S rRNA-Gene-Targeted Group-Specific Primers for the Detection and Identification of Predominant Bacteria in Human Feces. Appl. Environ. Microbiol. 68, 5445–5451. doi: 10.1128/AEM.68.11.5445-5451.2002
McLaren, M. R., Callahan, B. J. (2020). Pathogen Resistance may be the Principal Evolutionary Advantage Provided by the Microbiome. Philos. Trans. R. Soc Lond. B Biol. Sci. 375, 20190592. doi: 10.1098/rstb.2019.0592
Medeiros, A. A. (1972). Bacteroides Bacillemia. Sepsis for Surgeons. Arch. Surg. 105, 819–820. doi: 10.1001/archsurg.1972.04180120006001
Mitchell, C. M., Mazzoni, C., Hogstrom, L., Bryant, A., Bergerat, A., Cher, A., et al. (2020). Delivery Mode Affects Stability of Early Infant Gut Microbiota. Cell Rep. Med. 1, 100156. doi: 10.1016/j.xcrm.2020.100156
Palmer, C., Bik, E. M., DiGiulio, D. B., Relman, D. A., Brown, P. O. (2007). Development of the Human Infant Intestinal Microbiota. PloS Biol. 5, e177. doi: 10.1371/journal.pbio.0050177
Pasolli, E., Asnicar, F., Manara, S., Zolfo, M., Karcher, N., Armanini, F., et al. (2019). Extensive Unexplored Human Microbiome Diversity Revealed by Over 150,000 Genomes From Metagenomes Spanning Age, Geography, and Lifestyle. Cell 176, 649–662.e20. doi: 10.1016/j.cell.2019.01.001
Plaza-Díaz, J., Fontana, L., Gil, A. (2018). Human Milk Oligosaccharides and Immune System Development. Nutrients 10, 1038–1055. doi: 10.3390/nu10081038
Pricop, G. R., Gheorghe, I., Pircalabioru, G. G., Cristea, V., Popa, M., Marutescu, L., et al. (2020). Resistance and Virulence Features of Bacteroides Spp. Isolated From Abdominal Infections in Romanian Patients. Pathogens 9, 940–950. doi: 10.3390/pathogens9110940
Pruss, K. M., Marcobal, A., Southwick, A. M., Dahan, D., Smits, S. A., Ferreyra, J. A., et al. (2021). Mucin-Derived O-Glycans Supplemented to Diet Mitigate Diverse Microbiota Perturbations. ISME J. 15, 577–591. doi: 10.1038/s41396-020-00798-6
R Core Team (2021). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing (Vienna, Austria). Available at: https://www.R-project.org/.
Salli, K., Hirvonen, J., Siitonen, J., Ahonen, I., Anglenius, H., Maukonen, J. (2021). Selective Utilization of the Human Milk Oligosaccharides 2’-Fucosyllactose, 3-Fucosyllactose, and Difucosyllactose by Various Probiotic and Pathogenic Bacteria. J. Agric. Food Chem. 69, 170–182. doi: 10.1021/acs.jafc.0c06041
Salyers, A. A., Vercellotti, J. R., West, S. E., Wilkins, T. D. (1977). Fermentation of Mucin and Plant Polysaccharides by Strains of Bacteroides From the Human Colon. Appl. Environ. Microbiol. 33, 319–322. doi: 10.1128/aem.33.2.319-322.1977
Sela, D. A., Chapman, J., Adeuya, A., Kim, J. H., Chen, F., Whitehead, T. R., et al. (2008). The Genome Sequence of Bifidobacterium Longum Subsp. Infantis Reveals Adaptations for Milk Utilization Within the Infant Microbiome. Proc. Natl. Acad. Sci. U. S. A. 105, 18964–18969. doi: 10.1073/pnas.0809584105
Sela, D. A., Mills, D. A. (2010). Nursing Our Microbiota: Molecular Linkages Between Bifidobacteria and Milk Oligosaccharides. Trends Microbiol. 18, 298–307. doi: 10.1016/j.tim.2010.03.008
Shishkin, A. A., Giannoukos, G., Kucukural, A., Ciulla, D., Busby, M., Surka, C., et al. (2015). Simultaneous Generation of Many RNA-Seq Libraries in a Single Reaction. Nat. Methods 12, 323–325. doi: 10.1038/nmeth.3313
Smith-Brown, P., Morrison, M., Krause, L., Davies, P. S. W. (2016). Mothers Secretor Status Affects Development of Childrens Microbiota Composition and Function: A Pilot Study. PloS One 11, e0161211. doi: 10.1371/journal.pone.0161211
Sonnenburg, J. L., Xu, J., Leip, D. D., Chen, C.-H., Westover, B. P., Weatherford, J., et al. (2005). Glycan Foraging In Vivo by an Intestine-Adapted Bacterial Symbiont. Science 307, 1955–1959. doi: 10.1126/science.1109051
Sprouffske, K., Wagner, A. (2016). Growthcurver: An R Package for Obtaining Interpretable Metrics From Microbial Growth Curves. BMC Bioinf. 17, 172. doi: 10.1186/s12859-016-1016-7
Stewart, C. J., Ajami, N. J., O’Brien, J. L., Hutchinson, D. S., Smith, D. P., Wong, M. C., et al. (2018). Temporal Development of the Gut Microbiome in Early Childhood From the TEDDY Study. Nature 562, 583–588. doi: 10.1038/s41586-018-0617-x
Turroni, F., Duranti, S., Bottacini, F., Guglielmetti, S., Van Sinderen, D., Ventura, M. (2014). Bifidobacterium Bifidum as an Example of a Specialized Human Gut Commensal. Front. Microbiol. 5, 437. doi: 10.3389/fmicb.2014.00437
Vandenplas, Y., Berger, B., Carnielli, V. P., Ksiazyk, J., Lagström, H., Sanchez Luna, M., et al. (2018). Human Milk Oligosaccharides: 2’-Fucosyllactose (2'-FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients 10, 1161–1173. doi: 10.3390/nu10091161
Vatanen, T., Plichta, D. R., Somani, J., Münch, P. C., Arthur, T. D., Hall, A. B., et al. (2019). Genomic Variation and Strain-Specific Functional Adaptation in the Human Gut Microbiome During Early Life. Nat. Microbiol. 4, 470–479. doi: 10.1038/s41564-018-0321-5
Walsh, C., Lane, J. A., van Sinderen, D., Hickey, R. M. (2020). Human Milk Oligosaccharides: Shaping the Infant Gut Microbiota and Supporting Health. J. Funct. Foods 72, 104074. doi: 10.1016/j.jff.2020.104074
Wang, M., Li, M., Wu, S., Lebrilla, C. B., Chapkin, R. S., Ivanov, I., et al. (2015). Fecal Microbiota Composition of Breast-Fed Infants is Correlated With Human Milk Oligosaccharides Consumed. J. Pediatr. Gastroenterol. Nutr. 60, 825–833. doi: 10.1097/MPG.0000000000000752
Wexler, A. G., Goodman, A. L. (2017). An Insider’s Perspective: Bacteroides as a Window Into the Microbiome. Nat. Microbiol. 2, 17026. doi: 10.1038/nmicrobiol.2017.26
Xu, J., Bjursell, M. K., Himrod, J., Deng, S., Carmichael, L. K., Chiang, H. C., et al. (2003). A Genomic View of the Human-Bacteroides Thetaiotaomicron Symbiosis. Science 299, 2074–2076. doi: 10.1126/science.1080029
Yassour, M., Jason, E., Hogstrom, L. J., Arthur, T. D., Tripathi, S., Siljander, H., et al. (2018). Strain-Level Analysis of Mother-To-Child Bacterial Transmission During the First Few Months of Life. Cell Host Microbe 24, 146–154.e4. doi: 10.1016/j.chom.2018.06.007
Yassour, M., Vatanen, T., Siljander, H., Hämäläinen, A.-M., Härkönen, T., Ryhänen, S. J., et al. (2016). Natural History of the Infant Gut Microbiome and Impact of Antibiotic Treatment on Bacterial Strain Diversity and Stability. Sci. Transl. Med. 8, 343ra81. doi: 10.1126/scitranslmed.aad0917
Yatsunenko, T., Rey, F. E., Manary, M. J., Trehan, I., Dominguez-Bello, M. G., Contreras, M., et al. (2012). Human Gut Microbiome Viewed Across Age and Geography. Nature 486, 222–227. doi: 10.1038/nature11053
Zabel, B. E., Gerdes, S., Evans, K. C., Nedveck, D., Singles, S. K., Volk, B., et al. (2020). Strain-Specific Strategies of 2′-Fucosyllactose, 3-Fucosyllactose, and Difucosyllactose Assimilation by Bifidobacterium Longum Subsp. Infantis Bi-26 and ATCC 15697. Sci. Rep. 10, 15919–15937. doi: 10.1038/s41598-020-72792-z
Keywords: infant microbiome, human milk oligosaccharides (HMO), Bacteroides, microbial RNA seq, breastmilk
Citation: Kijner S, Cher A and Yassour M (2022) The Infant Gut Commensal Bacteroides dorei Presents a Generalized Transcriptional Response to Various Human Milk Oligosaccharides. Front. Cell. Infect. Microbiol. 12:854122. doi: 10.3389/fcimb.2022.854122
Received: 13 January 2022; Accepted: 22 February 2022;
Published: 18 March 2022.
Edited by:
Gislane Lelis Vilela de Oliveira, São Paulo State University, BrazilReviewed by:
Daniel A. Medina, San Sebastián University, ChileChristopher James Stewart, Newcastle University, United Kingdom
Sabrina Neves Casarotti, Federal University of Mato Grosso, Brazil
Copyright © 2022 Kijner, Cher and Yassour. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Moran Yassour, moranya@mail.huji.ac.il